Uptake of Sulfate from Ambient Water by Freshwater Animals


1
U.S. Environmental Protection Agency, Office of Research and Development, National Center for Environmental Assessment, Cincinnati, OH 45268, USA
2
U.S. Environmental Protection Agency, Office of Research and Development, Center for Environmental Measurement and Modeling, Cincinnati, OH 45268, USA
3
U.S. Environmental Protection Agency, Office of Research and Development, National Exposure Research Laboratory, Cincinnati, OH 45268, USA
4
Pegasus Technical Services, Inc., Cincinnati, OH 45268, USA
*
Author to whom correspondence should be addressed.
Water 2020, 12(5), 1496; https://doi.org/10.3390/w12051496
Submission received: 3 April 2020
/
Revised: 12 May 2020
/
Accepted: 20 May 2020
/
Published: 23 May 2020
(This article belongs to the Special Issue Applied Ecology Research for Water Quality Management)
Abstract
:To better understand how the sulfate (SO42−) anion may contribute to the adverse effects associated with elevated ionic strength or salinity in freshwaters, we measured the uptake and efflux of SO42− in four freshwater species: the fathead minnow (Pimephales promelas, Teleostei: Cyprinidae), paper pondshell (Utterbackia imbecillis, Bivalvia: Unionidae), red swamp crayfish (Procambarus clarkii, Crustacea: Cambaridae), and two-lined mayfly (Hexagenia bilineata, Insecta: Ephemeridae). Using δ(34S/32S) stable isotope ratios and the concentrations of S and SO42−, we measured the SO42− influx rate (Jin), net flux (Jnet), and efflux rate (Jout) during a 24 h exposure period. For all four species, the means of Jin for SO42− were positive, and Jin was significantly greater than 0 at both target SO42− concentrations in the fish and mollusk and at the lower SO42− concentration in the crayfish. The means of Jout and Jnet were much more variable than those for Jin, but several species by target SO42− concentration combinations for Jout and Jnet, were negative, which suggests the net excretion of SO42− by the animals. The results of our experiments suggest a greater regulation of SO42− in freshwater animals than has been previously reported.
1. Introduction
Water quality benchmarks were recently developed for elevated total ion concentrations in freshwaters using specific conductance as a measurement endpoint to protect aquatic life [1,2]. However, there is still uncertainty about how different ions contribute to the adverse effects associated with elevated total ionic strength or salinity on freshwater biota, and some studies have suggested a more traditional approach where the concentrations of specific anions should be the targets of chemical monitoring and ambient water quality criteria [3,4,5,6]. The predominant form of dissolved sulfur (S) in water is the anion, sulfate (SO42−), which can be especially elevated in waters affected by mining. Mining exposes sulfide-rich minerals, such as pyrite associated with coal mining and other metal sulfides associated with hard rock metal mining [7,8,9,10]. Under oxic conditions, these sulfides are rapidly transformed into SO42−.
Several studies have recently attempted to assess the ecotoxicity of SO42− with standard bioassays that have exclusively used Na2SO4 [11,12,13,14]. The objective of at least some of these studies is the compilation of sufficient bioassay data for a species sensitivity distribution to develop an ambient water criterion for SO42−. These studies attribute many of the observed adverse effects to SO42−. This assumes that any effects associated with the concurrently elevated concentrations of Na+ are minor, even though the uptake of Na+ across epithelial membranes is well known [15,16,17]. Additionally, the uptake of SO42− across external epithelial membranes, such as gills, occurs and has an ionoregulatory or osmoregulatory adverse effect on these animals. However, because the transport physiology of SO42− in freshwater animals is relatively unstudied, the validity of the second assumption is unknown. Other studies have begun to study the interactions between different anion and cation combinations [18,19,20].
To support decisions relating to the potential for adverse effects associated with individual major ions, Na+, K+, Ca2+, Mg2+, Cl−, SO42−, and HCO3−, we previously published a literature review on the ion physiology of freshwater species of four animal groups: teleost fish, crustaceans, aquatic insects, and mollusks [21]. While this review found extensive literature on many of these major ions at least for teleost fish, if not always for the freshwater invertebrates, it identified a data gap related to the ionoregulatory physiology of SO42−. Limited data appear to suggest that SO42− is relatively impermeant to the gill membranes of fish [22,23], crustaceans [24,25], or unionid mussels [26]. However, SO42−-transporters are present in amphibian skin [27], which functions in ion uptake similarly to the gill membranes of fish, crustaceans, and mollusks or to anal papillae, chloride epithelia, or other epithelial surfaces in aquatic insects [21]. However, see the research on SO42− uptake from the water that has been recently published on aquatic insects by Scheibener et al. [28] and Buchwalter et al. [29].
Like other major ions, SO42− is present in the extracellular fluids (i.e., blood or hemolymph) of freshwater animals. Reported SO42− concentrations include 0.76 ± 0.18 mmol/L (n = 6); 2.58 ± 0.06 (n = 30) and 2.60 ± 0.90 mmol/L (n = 7) in two species of Anodonta and zebra mussels (Dreissena polymorpha), respectively (Mollusca) [30,31,32]; 0.94 ± 0.26 for the spinycheek crayfish (Orconectes limosus) [33]; and 0.89 and 2.14 mmol/L for white sucker (Catostomus commersoni) and rainbow trout (Oncorhynchus mykiss), respectively [34]. A greater SO42− concentration of 18.7 mmol/L was observed in freshwater Japanese eels (Anguilla japonica) [35], but the authors suggest this catadromous species balances greater SO42− with less Cl− in its blood.
Moreover, SO42− is metabolically active. S is a component of the amino acids methionine and cysteine, which produce S-containing proteins, and a component of other biomolecules, such as sulfated glycosaminoglycans, which are generally ubiquitous in metazoans, including mollusks, crustaceans, insects, and fish [36,37,38,39]. In most organisms, the initial step in S assimilation is SO42− activation to 3‘-phosphoadenosine 5’-phosphosulfate (PAPS) [40,41,42,43].
Freshwater animals are generally hyperregulators that maintain greater ion concentrations in their blood or hemolymph than are found in surrounding freshwaters [44]. As the external medium is hypoosmotic or more dilute than body fluids, these species deal with the continuous diffusional loss of salts and the osmosis of water across their permeable membranes. Therefore, water balance is accomplished by the excretion of dilute waste fluids by their renal systems. Salt concentrations are maintained by the function of various ion transporting proteins in epithelial membranes, such as in the gills, gastrointestinal system, or renal system, that allow the active transport of ions against concentration gradients (i.e., the absorption of ions from the surrounding water or food or the reabsorption of ions from waste fluids, such as urine). Furthermore, most freshwater species, unlike saltwater species, limit their drinking of water, thereby limiting the absorption of water through the gastrointestinal system and dilution of the hemolymph. However, increased water concentrations of some ions, such as SO42−, change concentration gradients across the epithelial membranes involved in ionoregulation, such as the gills. This may change SO42− influx across these membranes, increase blood or hemolymph SO42− concentrations, and have osmoregulatory or other adverse effects. However, it is also possible that these epithelial membranes could be impermeant to SO42−, and none of the above effects may occur. To test whether SO42− can move across epithelial membranes, we conducted laboratory experiments with a fish, a crustacean, a mollusk, and a mayfly. These experiments used protocols similar to traditional toxicity tests in that the organisms were exposed to reconstituted water with elevated SO42− concentrations. However, the concentrations were not expected to have overtly adverse effects, such as mortality. Moreover, the reconstituted water was made, in part, with enriched Na2[34S]O4 to elevate its δ(34S/32S). After exposure, the whole body stable isotope ratios of S were measured to assess whether the 34S associated with animal tissues increased.
This study was designed to fill a data gap by conducting laboratory experiments with species from four freshwater animal groups (crustaceans, fish, unionid mussels, and aquatic insects) to measure SO42− uptake from ambient waters and the effect of ambient SO42− concentrations on SO42− uptake. Significant SO42− uptake from the water into an animal would show that the epithelial membranes are permeant to SO42−, likely via an ion-specific transporter. This SO42− uptake would affect internal SO42− concentrations, would play a direct role in ionoregulation by the animal, and could have adverse effects if elevated in freshwaters.
2. Materials and Methods
To test whether SO42− uptake is similar or differs among different major taxa groups of freshwater animals, we conducted parallel experiments using a representative species in each of four taxa groups. The ion transport physiology of these species is unlikely to change with the aquatic developmental stage [21]. Therefore, juvenile fish and crayfish, adult unionid mussels, and later instar mayfly nymphs were used in the experiments. The exposures for each of the four species was conducted during a single 24 h period, but the experiments were conducted at different times between September 2017 and April 2019.
2.1. Test Animals
Although we considered, originally, using model species (e.g., Cladocera and Chironomidae), often used by toxicity studies, our methods required larger individuals to provide sufficient biomass for stable isotope analysis. We selected the fathead minnow (Pimephales promelas Rafinesque, 1820) which is commonly used as a model for a teleost fish. Less conventional test species included a mollusk, the paper pondshell (Utterbackia imbecillis (Say, 1829)); a crustacean, the red swamp crayfish (Procambarus clarkii (Girard, 1852)); and an aquatic insect, the two-lined mayfly (Hexagenia bilineata (Say, 1824)).
We used fathead minnows from laboratory colonies usually cultured and maintained for toxicity testing according to [45,46]. We fed them live Artemia nauplii twice a day at a rate of 1 mL per 20 L tank per day-of-age per feeding with a nauplii suspension of 15 mL of brine shrimp cysts (Brine Shrimp Direct, Ogden, UT, USA) incubated for 24 h at 28 °C in aerated Labline water with 25 mL NaCl added.
We purchased red swamp crayfish from Carolina Biological Supply Co., Burlington, NC, USA. The crayfish were placed in a tank with about 175 L of water aerated with air stones. Lengths of polyvinyl chloride pipe cut in half lengthwise were placed in the aquaria to provide cover for the crayfish and reduce aggression. The crayfish were fed thawed adult Artemia ad libitum daily.
We obtained the paper pondshells from the Kentucky Department of Fish and Wildlife Resources’ Center for Mollusk Conservation (Frankfort, KY, USA). The mussels were held in aquaria, with 28 individuals per aquarium, where the water was aerated by an air stone. The mollusks were fed 20 mL of FFAY, an internally made mixture of fish flake food (Tetramin®, Tetra, Blacksburg, VA, USA), alfalfa (from capsule; Nature’s Way, Fargo, ND, USA), and yeast (Flieschmann’s, Oakbrook Terrace, IL, USA); and 15 mL of an algal culture of flagellated algae and diatoms (Shellfish Diet 1800®, Reed Mariculture, Campbell, CA, USA) and 15 mL of alfalfa per tank daily.
We purchased the two-lined mayfly from The Reel Thing Live Bait, Green Bay, WI, USA. The mayfly nymphs were held in aquaria, where the water was aerated by an air stone, and were fed ground cereal grain flake food (Cerophyll®, Ward’s Natural Science Establishment, Inc., Rochester, NY, USA).
Before their use in experiments, we held the fish and invertebrates in dechlorinated and hardness-adjusted municipal tap water (mean ionic composition in mmol L−1: 1.04 [Na+], 0.05 [K+], ~0.25–0.55 [Ca2+], 0.43 [Mg2+], ≤ 0.01 [Cl−], and 0.93 [SO42−]; pH: 7.2–8.1; total organic carbon (TOC): ~0.08 mg C L−1; and hardness: ~150–180 mg L−1 as CaCO3). All animals, except the two-lined mayflies, were maintained at a constant temperature of 25 ± 1 °C with a 16:8 h light to dark photoperiod. The two-lined mayflies were maintained at a constant temperature of 15 ± 1 °C.
For at least 24 h before the individual experiments, the animals were acclimated to a modified moderately hard reconstituted water (MMHRW) similar to the R-MHRW of Smith et al. [47] generated by the addition of reagent-grade salts to deionized water produced by a Millipore Super-Q Plus water purification system and bubbled with CO2 to dissolve the CaCO3, particularly using Na2SO4 (Table 1). During the 24 h acclimation and test phases, the animals were not fed, while the holding water temperatures were maintained.
2.2. Test Water
Upon the initiation of the experiment, the animals were exposed to two concentrations of SO42−, with the MMHRW with Na2SO4 added to increase by 2× (i.e., 0.49 mmol L−1) or 5× (1.23 mmol L−1) the SO42− concentrations to test whether differing water SO42− concentrations influenced uptake (Table 1). This addition also increased Na+ concentrations to 1.03 mmol L−1 and 2.51 mmol L−1, respectively. None of these concentrations are near the concentrations that have been observed to have acute adverse effects on freshwater animals [6,11,12,48]. Moreover, the Na+/K+ molar ratios were 22.0 and 53.5, respectively, which are within the range where alterations of this ratio have not been observed to have osmoregulatory effects [49,50].
To observe influx rates, test water made with unenriched reagent grade Na2SO4 was mixed with the same concentration test water made with 90% atom enriched Na2[34S]O4 (Sigma-Aldrich, Miamisburg, OH, USA) at a ratio of 939 parts of reagent grade test water to 61 parts of 90% atom enriched test water. A test water made with only 90% atom enriched Na2[34S]O4 could not be used without dilution to reduce the enrichment of the 34S. Too strong of a signal from 34S when the samples are analyzed in the mass spectrometer can overload the detectors and cause falsely elevated concentration readings in subsequent samples (R. Venkatapathy, personal communication).
2.3. Sampling Methods
2.3.1. Measurements of Animals and Water
Measurements were made on organisms placed in containers with aeration, containing 200 mL, 12 L, 7 L, and 150 mL of the waters for the minnow, unionid mollusk, crayfish, and mayfly, respectively. The volumes were chosen based on the mass loading limits for the animals in static tests [45]. The two-lined mayflies were supplied with short lengths (~38 mm) of 9.5 mm inner-diameter vinyl tubing as artificial burrows [51]. In all four species, the mass of the animal (i.e., ≥10 mg dry mass) was sufficient for chemical analysis, permitting us to measure each individual.
Subsamples of the test waters were taken to measure the SO42− concentrations and ẟ(34S/32S) for each exposure at 0 h, and a water sample was taken from each exposure container to measure the SO42− concentrations after the 24 h exposures. At least 10 extra individuals of each species were sacrificed at 0 h to measure initial whole-body S concentrations and ẟ(34S/32S). Measurements from 25 separate replicates of each SO42− concentration were collected for each test species. From every exposure, replicate whole-body samples of each test species were taken at the end of the exposure to the enriched stable isotope water and used to measure whole-body S concentrations and ẟ(34S/32S). However, because there was some mortality in the two-lined mayfly experiment, the numbers of replicates were 17 and 22 for the 0.49 and 1.25 mmol L−1 SO42− exposures, respectively. Moreover, an error in the initial water SO42− analysis in the mayfly experiment reduced the number of valid measurements of the initial 0.49 mmol L−1 water SO42− concentration to 1, precluding valid means of Jout and Jnet for the 0.49 mmol L−1 SO42− exposure.
At the end of each exposure, the fish, crayfish, or mayfly nymph individuals were removed from the containers, rinsed in deionized water to remove any stable isotope label from the surface, and blotted dry on filter paper. For the unionid mussel, the soft tissue was cut from the shell. The fish, mussels (soft tissue only), and mayfly nymphs were dried overnight at 90 °C, whereas the crayfish were freeze-dried for 14 days to weaken their exoskeletons. The animals were then weighed, homogenized, and powdered to 100–200 μm with a mortar and pestle, stored in vials, and analyzed for total S and δ(34S/32S) [52].
2.3.2. Chemical Analysis of Test Water and Animals
The total concentrations of SO42− in the test waters were measured with ion chromatography (EPA Method 300.0) [53]. For the stable isotope analysis of SO42− in the initial test waters, we precipitated the dissolved SO42− in subsamples of the water placed in the test chambers as barium sulfate (BaSO4) following Révész et al. [54].
The total S concentration of the dried animal tissues was measured using an elemental analyzer. The molar ratios of 34S/32S, reported as the deviation of this ratio from the international standard, Canon Diablo Troilite, or δ(34S/32S) [55], were measured in both the BaSO4 precipitates and dried animal tissues with an elemental analyzer connected to a continuous-flow 20-20 gas source stable isotope ratio mass spectrometer [52,54]. For 34S analyses, the sample was placed in a tin capsule with vanadium dioxide for combustion, the combustion reactor was held at 1080 °C, and the reaction tube contained tungsten oxide on alumina as an oxidative catalyst and copper metal to remove excess O2 subsequent to combustion. The S in the sample was converted to SO2 and, along with N2 and CO2, was passed through two H2O traps. The purified gases were then separated in a 30 cm, 0.5” OD, QS GC column held at 45 °C and passed into the mass spectrometer—N2 and CO2 first, and then SO2. Approximately one-third of the SO2 was cracked to SO in the source, allowing 34S to be measured by continuously monitoring masses 48, 49, and 50. The mass peaks were plotted, the area under each mass peak was determined, and the isotope ratios were calculated. These ratios were referenced to ratios determined on in-house reference materials analyzed in the same analytical run. The raw ratio data were corrected for drift over the course of the run along with blank/linearity effects, and if present, then normalized to the international standards. Analytical precisions, based on the replicate analyses of international reference materials, were ±0.3‰ for δ34S.
2.3.3. Compartmental Analysis to Calculate the Influx and Efflux of Sulfate
The SO42−, total S, and δ34S ratio data were used in a compartmental analysis of a single pool system for using stable isotope tracer data [56,57] modified from models using radioisotope data for ion uptake by fish and crayfish [58,59,60]. Compartmental analysis models the movement of a solute between two compartments based on diffusion and mass conservation. In this system, we are measuring the movement of SO42− between the surrounding water and the animal across semipermeable epithelial cell membranes that may be facilitated by the presence of SO42−-specific transporter proteins. This is because the SO42− in the test water can be traced with the artificially elevated levels of 34S by the SO42−-influx rate (Jin). However, because only the change in the SO42− concentration in the test waters was used to measure the net SO42−-flux rate (Jnet), the SO42−-efflux rate (Jout) may include S from sources other than the test water, such as food.
2.3.4. Calculations
Jnet was measured as the change in the water SO42− concentration during the exposure period (t = 1 day):
where [SO42−]0 and [SO42−]t are the concentrations of SO42− in the water (μmol L−1) at the beginning and end of the exposure period, respectively; V is the volume of the water in the chamber (L) measured at t = 0 and t ≈ 24 h to account for evaporation; M is the mass (g) of the animal placed into the container; and t is the length of the exposure period (days).
Jin was measured as the changes in the fractional molar abundance of 34S and total S concentration in the animal during the exposure interval relative to the initial fractional molar abundance of 34S in the test waters:
where X(34S)int(0) is the initial fractional molar abundance of 34S in the animal, [S]int(0) is the initial concentration of S in the animal (μmol g−1), X(34S)int(t) is the fractional molar abundance of 34S at the end of the exposure, [S]int(t) is the concentration of S in the animal at the end of the exposure (μmol /g−1), X(34S)bath is the fractional molar abundance of 34S in the test waters, and t is the length of the exposure period (day).
The fractional molar abundance of 34S or X(34S) for a sample is calculated from the molar ratio of 34S to 32S or R(34S/32S) for the sample by:
and R(34S/32S) is calculated from the reported δ(34S/32S) by:
where N(34S)std/N(32S)std is the molar ratio of the heavy stable isotope in the standard material, Canyon Diablo Troilite, which by convention is assigned a N(34S)/N(32S) of 0.045005 [61]. The values are divided by 1000, because δ(34S/32S) is reported in parts per mille relative to the standard material.
2.3.5. Statistical Analysis
As the measurements of SO42− and δ(34S/32S) in the water and in animals at the beginning of the exposures were made on and summarized for replicate subsamples, the variation associated with these measurements was pooled as appropriate with the variation of the calculated variables, Jin, Jout, and Jnet. Then, the calculated variables for each species were tested to determine whether each variable was significantly different from 0 using a t-test (PROC TTEST, SAS Institute, Cary, NC, USA). Because six variable-by-concentration combinations were tested for each species, a Bonferroni adjustment of p = 0.0083 was used.
3. Results
The measured SO42− concentrations in the artificial water used with the different species were variable compared to the target concentrations (Table 2). The measured δ(34S/32S) for the artificial waters (Table 2) were more than 100 times the initial measured δ(34S/32S) of the animal tissues, which were +11.386 ± 0.090, −1.420 ± 0.910, −2.304 ± 0.987, and −4.617 ± 0.530 for the fathead minnows, paper pondshells, red swamp crayfish, and two-lined mayflies, respectively. The summary statistics for all the variables used in the compartmental analyses may be found in Table S1.
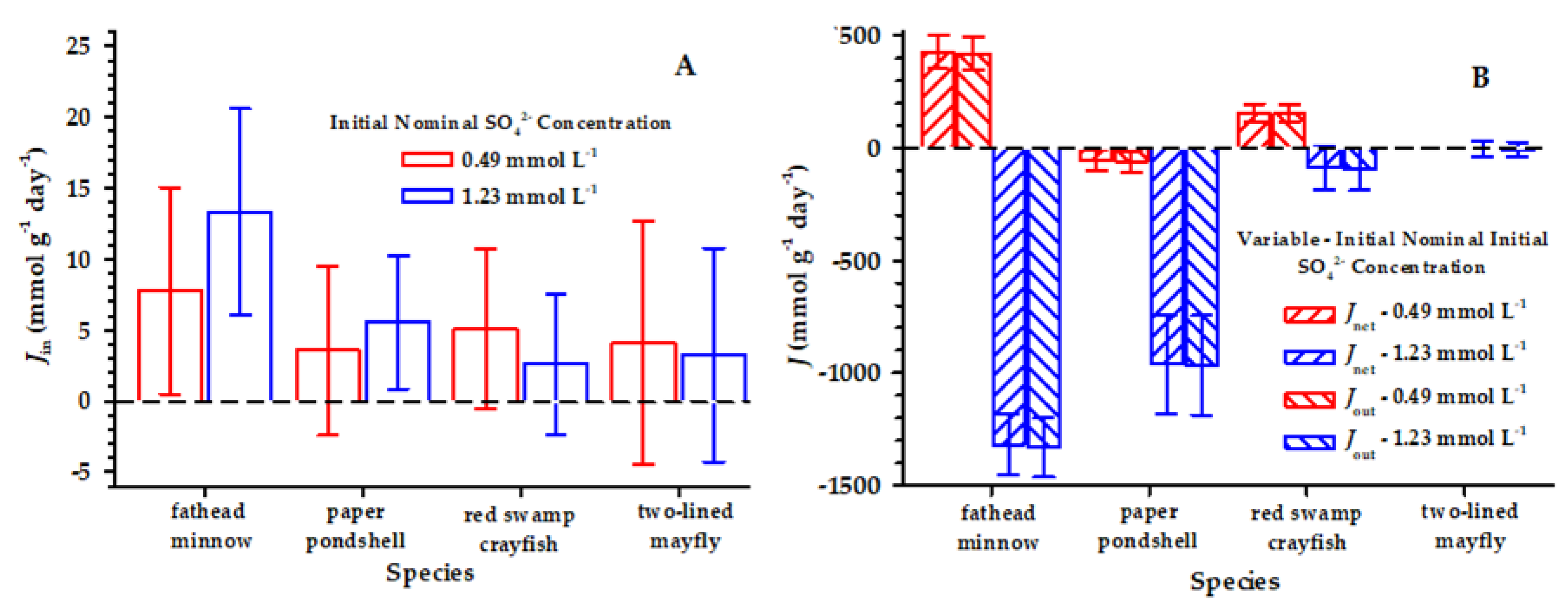
The means of Jin for SO42− were positive for all the species and ranged from 2.14 to 13.32 μmol g−1 day−1 among the species and two nominal sulfate concentrations (Figure 1). The Jin in 5 of 50, 12 of 50, 13 of 50, and 8 of 39 individual exposures of the fish, mollusk, crayfish, and mayfly were negative, and there were large confidence bounds around the means (i.e., the coefficient of variation ranges from 0.54 to 3.63). In part, notable variance was added to the means of Jin because of unexpected variation in the measurements of the initial animal S concentrations (Table 3), but Jin was significantly greater than 0 at both target SO42− concentrations in the fish and mollusk and at the lower SO42− concentration (i.e., 0.49 mmol L−1) in the crayfish (Table 4). Additionally, Jin increased between the two SO42− concentrations in the fish and mollusk (Figure 1), although the increase was statistically significant only for the fish (df = 48, t = 2.70, p = 0.009) and the mollusk (df = 48, t = 1.13, p = 0.20).
4. Discussion
Because we measured an increase in whole animal R(34S/32S) in animals exposed to water where the added SO42− was highly enriched with 34S, Jin measured the uptake of SO42− from the water. Presumably, the SO42− moved through SO42−-transporters on external epithelial membranes, such as the gills, chloride cells, or integument, because other ions commonly move through epithelial membranes via various intercellular pathways involving various ion transporters. Although our methods used whole animal assays and therefore did not definitively distinguish between external surfaces and internal tissues, the measured Jin, when expressed in the same units, are of a similar range of magnitude as the measurements of the uptake of other ions, including Cl− anions, at similar water concentrations [62,63,64,65]. Even though our methods do not distinguish between ionocyte-mediated transport and paracellular transport, which has been described for other ions, paracellular transport does not generally occur against concentration gradients, and we expect that it would take longer than 24 h for the equilibration of the isotope between the test solution and the internal milieu without the aid of facilitated or active transport [66,67,68,69].
The larger Jout suggests that there is an internal pool of SO42− supplied by sources in addition to uptake from the water, such as from food [70,71]. By measuring the change in the SO42− concentration in the water, Jout and Jnet measure all the effluxes of SO42− between the animals and the water, including renal excretion.
A number of other studies have observed SO42− transporters in aquatic animals, primarily associated with internal epithelial membranes. In teleost fish, two types of epithelial SO42− transporters, a Na+/SO42−-cotransporter (NaS1, SLC13 family) and a SO42−/anion-exchanger (Sat1, SLC26 family), have been identified in the proximal tubules of the kidneys that are primarily involved in SO42− excretion and resorption [35,72,73]. The functional analysis of freshwater mollusks suggests a similar renal regulation of SO42− [26,30]. In mosquito (Aedes campestris) larvae from saline lakes, transporters associated with the Malpighian tubules excrete SO42− [74,75]. In marine Atlantic lobsters (Homarus americanus), SO42− transporters associated with hepatopancreatic epithelia excrete SO42− in exchange for either C2H42− or Cl− [76,77,78,79]. The Na+/SO42−-cotransporter, NaS1, was also detected in the intestinal tissue of zebrafish (Danio rerio) but not gill tissues [72]. However, our experiments were not designed to identify the types of SO42− transporters beyond their role in uptake from the ambient water.
The amino acid cysteine and S-containing proteins are synthesized in metazoans from the amino acid methionine. While the initial step is SO42− activation to PAPS, S is added to methionine by the sulfate assimilatory reduction of PAPS, a pathway not found in metazoans [40,41,42,43]. Therefore, these amino acids are sources of S in animals via ingestion. In other biomolecules, sulfate groups are transferred from PAPS by a sulfonation pathway, which is found in metazoans [41,43]. Therefore, inorganic SO42− from some source is required by animals. The renal reabsorption of SO42− is likely part of the source, but the source is also supplied by the direct uptake of SO42− from the water, which we observed in our experiments.
In freshwater nymphs of Plecoptera, Ephemeroptera, and Trichoptera, Scheibener et al. [28] measured the uptake of SO42− and found that this uptake was inhibited by increased Na+ water concentrations, suggesting the presence of a Na+/SO42−-cotransporter. Buchwalter et al. [29] identified similar SO42− transporters in the mayfly, Neocloeon trangulifer, but the localization of these transporters was not determined. This research also observed that SO42− uptake increased with an increasing SO42− water concentration.
In conclusion, our study, along with studies from the Buchwalter laboratory [28,29], suggests that there is direct uptake of SO42− from the water in these four groups of freshwater animals. Additionally, there is some evidence that this uptake may increase with the water concentration of SO42−. However, the uptake of SO42− from the water is not the only source of S, and S from food likely contributes to the SO42− excreted by these animals. Therefore, elevated water SO42− may interact with other ions to have ionoregulatory effects in freshwater animals that could cause the effects observed by more traditional ecotoxicological studies.
A next step for research on any of these freshwater animals would be to sequence, locate, and functionally characterize any SO42−-transporters on their gills or other external epithelial membranes, as has been done for other ions [80,81,82,83]. Such information would clarify the potential interactions between SO42− and other ions, such as Na+, when these ions are elevated in freshwaters. This will more completely identify the potential pathways for adverse outcomes [84] for elevated SO42− in freshwaters and better support risk assessments, leading to the development of water-quality benchmarks or criteria.
Supplementary Materials
The following are available online at https://www.mdpi.com/2073-4441/12/5/1496/s1, Table S1: Summary statistics (mean, standard error (SE), and n for all variables used in the equations to estimate Jin, Jout, and Jnet (μM g−1 day−1).
Author Contributions
Conceptualization, M.B.G. and J.M.L.; methodology, M.B.G., J.M.L. and H.H.; validation, M.B.G. and H.H.; formal analysis, M.B.G.; investigation, M.B.G., J.M.L. and H.H.; resources, M.B.G. and J.M.L.; data curation, M.B.G. and H.H.; writing—original draft preparation, M.B.G. and H.H.; writing—review and editing, M.B.G. and J.M.L.; visualization, M.B.G. and supervision, M.B.G. and J.M.L.; All authors have read and agreed to the published version of the manuscript.
Funding
This study is based on work supported by the USEPA.
Acknowledgments
M. A. McGregor and staff of the Kentucky Department of Fish and Wildlife Resources’ Center for Mollusk Conservation (Frankfort, KY) cultured and supplied us with the paper floater adults. W. Thoeny (Pegasus Technical Services, Cincinnati, OH) assisted in the laboratory. The modification of the calculations for use with sulfur stable isotope data were reviewed by C. Wood (University of British Columbia, Vancouver, BC, Canada). Earlier drafts of this manuscript were reviewed and improved by comments from K. Fritz, S. Cormier (U.S. Environmental Protection Agency, Office of Research and Development, Cincinnati, OH) and three anonymous peer reviewers. This manuscript was prepared by USEPA, ORD, National Center for Environmental Assessment, Cincinnati Division and National Exposure Research Laboratory, Systems Exposure Division. It has been subjected to the agency’s peer and administration review and approved for publication. However, the views expressed are those of the authors and do not necessarily represent the views or policies of the USEPA. Any use of trade, firm, or product names is for descriptive purposes only and does not imply endorsement by the USEPA.
Conflicts of Interest
The authors declare no conflicts of interest.
Availability of Data
The compiled data set used in these analyses can be viewed or downloaded from USEPA’s Science Hub at https://doi.org/10.23719/1504445.
References
- Griffith, M.; Zheng, L.; Cormier, S.M. Using extirpation to evaluate ionic tolerance of freshwater fish. Environ. Toxicol. Chem. 2017, 37, 871–883. [Google Scholar] [CrossRef] [PubMed]
- Cormier, S.M.; Suter, G.; Zheng, L. Derivation of a benchmark for freshwater ionic strength. Environ. Toxicol. Chem. 2012, 32, 263–271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elphick, J.R.; Bergh, K.D.; Bailey, H.C. Chronic toxicity of chloride to freshwater species: Effects of hardness and implications for water quality guidelines. Environ. Toxicol. Chem. 2010, 30, 239–246. [Google Scholar] [CrossRef] [PubMed]
- Lasier, P.J.; Hardin, I.R. Observed and predicted reproduction ofCeriodaphnia dubiaexposed to chloride, sulfate, and bicarbonate. Environ. Toxicol. Chem. 2010, 29, 347–358. [Google Scholar] [CrossRef] [PubMed]
- Farag, A.M.; Harper, D.D. The chronic toxicity of sodium bicarbonate, a major component of coal bed natural gas produced waters. Environ. Toxicol. Chem. 2014, 33, 532–540. [Google Scholar] [CrossRef]
- Soucek, D.J.; Linton, T.K.; Tarr, C.D.; Dickinson, A.; Wickramanayake, N.; Delos, C.G.; Cruz, L.A. Influence of water hardness and sulfate on the acute toxicity of chloride to sensitive freshwater invertebrates. Environ. Toxicol. Chem. 2011, 30, 930–938. [Google Scholar] [CrossRef]
- Griffith, M.; Norton, S.B.; Alexander, L.C.; Pollard, A.I.; LeDuc, S.D. The effects of mountaintop mines and valley fills on the physicochemical quality of stream ecosystems in the central Appalachians: A review. Sci. Total Environ. 2012, 417, 1–12. [Google Scholar] [CrossRef]
- Cowie, R.; Williams, M.W.; Wireman, M.; Runkel, R.L. Use of Natural and Applied Tracers to Guide Targeted Remediation Efforts in an Acid Mine Drainage System, Colorado Rockies, USA. Water 2014, 6, 745–777. [Google Scholar] [CrossRef] [Green Version]
- Nordstrom, D.K. Hydrogeochemical processes governing the origin, transport and fate of major and trace elements from mine wastes and mineralized rock to surface waters. Appl. Geochem. 2011, 26, 1777–1791. [Google Scholar] [CrossRef]
- Iii, C.A.C.; Brightbill, R.A.; Langland, M.J. Abandoned Mine Drainage in the Swatara Creek Basin, Southern Anthracite Coalfield, Pennsylvania, USA: 1. Stream Water Quality Trends Coinciding with the Return of Fish. Mine Water Environ. 2010, 29, 176–199. [Google Scholar] [CrossRef] [Green Version]
- Elphick, J.R.; Davies, M.; Gilron, G.; Canaria, E.C.; Lo, B.; Bailey, H.C. An aquatic toxicological evaluation of sulfate: The case for considering hardness as a modifying factor in setting water quality guidelines. Environ. Toxicol. Chem. 2010, 30, 247–253. [Google Scholar] [CrossRef] [PubMed]
- Davies, T.D.; Hall, K.J. Importance of calcium in modifying the acute toxicity of sodium sulphate to Hyalella azteca and Daphnia magna. Environ. Toxicol. Chem. 2007, 26, 1243–1247. [Google Scholar] [CrossRef] [PubMed]
- Soucek, D.J. Comparison of hardness- and chloride-regulated acute effects of sodium sulfate on two freshwater crustaceans. Environ. Toxicol. Chem. 2007, 26, 773–779. [Google Scholar] [CrossRef] [PubMed]
- Soucek, D.J.; Dickinson, A. Full-life chronic toxicity of sodium salts to the mayfly Neocloeon triangulifer in tests with laboratory cultured food. Environ. Toxicol. Chem. 2015, 34, 2126–2137. [Google Scholar] [CrossRef] [PubMed]
- Horisberger, J.-D. Recent Insights into the Structure and Mechanism of the Sodium Pump. Physiology 2004, 19, 377–387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivanis, G.; Esbaugh, A.J.; Perry, S.F. Branchial expression and localization of SLC9A2 and SLC9A3 sodium/hydrogen exchangers and their possible role in acid-base regulation in freshwater rainbow trout (Oncorhynchus mykiss). J. Exp. Boil. 2008, 211, 2467–2477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pullikuth, A.; Aimanova, K.; Kang’Ethe, W.; Sanders, H.R.; Gill, S.S. Molecular characterization of sodium/proton exchanger 3 (NHE3) from the yellow fever vector, Aedes aegypti. J. Exp. Boil. 2006, 209, 3529–3544. [Google Scholar] [CrossRef] [Green Version]
- Soucek, D.J.; Mount, D.R.; Dickinson, A.; Hockett, J.R. Influence of dilution water ionic composition on acute major ion toxicity to the mayfly Neocloeon triangulifer. Environ. Toxicol. Chem. 2018, 37, 1330–1339. [Google Scholar] [CrossRef]
- Erickson, R.J.; Mount, D.R.; Highland, T.L.; Hockett, J.R.; Hoff, D.J.; Jenson, C.T.; Norberg-King, T.J.; Peterson, K.N. The acute toxicity of major ion salts to Ceriodaphnia dubia. II. Empirical relationships in binary salt mixtures. Environ. Toxicol. Chem. 2016, 36, 1525–1537. [Google Scholar] [CrossRef]
- Mount, D.R.; Erickson, R.J.; Highland, T.L.; Hockett, J.R.; Hoff, D.J.; Jenson, C.T.; Norberg-King, T.J.; Peterson, K.N.; Polaske, Z.M.; Wisniewski, S.; et al. The acute toxicity of major ion salts to Ceriodaphnia dubia: I. influence of background water chemistry. Environ. Toxicol. Chem. 2016, 35, 3039–3057. [Google Scholar] [CrossRef]
- Griffith, M. Toxicological perspective on the osmoregulation and ionoregulation physiology of major ions by freshwater animals: Teleost fish, crustacea, aquatic insects, and Mollusca. Environ. Toxicol. Chem. 2016, 36, 576–600. [Google Scholar] [CrossRef] [PubMed]
- Romeu, F.G.; Maetz, J. The Mechanism of Sodium and Chloride Uptake by the Gills of a Fresh-Water Fish, Carassius auratus. J. Gen. Physiol. 1964, 47, 1195–1207. [Google Scholar] [CrossRef] [PubMed]
- De Renzis, G.; Maetz, J. Studies on the mechanism of chloride absorption by the goldfish gill: Relation with acid-base regulation. J. Exp. Biol. 1973, 59, 339–358. [Google Scholar]
- Shaw, J. The absorption of sodium ions by the crayfish, Astacus pallipes Lereboullet: II. The effect of the external anion. J. Exp. Biol. 1960, 37, 534–547. [Google Scholar]
- McMahon, B.; Stuart, S. The physiological problems of crayfish in acid waters. In Acid Toxicity and Aquatic Animals; Cambridge University Press (CUP): Cambridge, UK, 1989; Volume 34, pp. 171–200. [Google Scholar]
- Dietz, T.H.; Udoetok, A.S.; Cherry, J.S.; Byrne, H.S.A. Kidney function and sulfate uptake and loss in the freshwater bivalve Toxolasma texasensis. Boil. Bull. 2000, 199, 14–20. [Google Scholar] [CrossRef]
- Larsen, E.H.; Simonsen, K. Sulfate transport in toad skin: Evidence for mitochondria-rich cell pathways in common with halide ions. Comp. Biochem. Physiol. Part A Physiol. 1988, 90, 709–714. [Google Scholar] [CrossRef]
- Scheibener, S.; Conley, J.M.; Buchwalter, D.B. Sulfate transport kinetics and toxicity are modulated by sodium in aquatic insects. Aquat. Toxicol. 2017, 190, 62–69. [Google Scholar] [CrossRef]
- Buchwalter, D.B.; Scheibener, S.; Chou, H.; Soucek, D.; Elphick, J. Are sulfate effects in the mayfly Neocloeon triangulifer driven by the cost of ion regulation? Philos. Trans. R. Soc. B Boil. Sci. 2018, 374, 20180013. [Google Scholar] [CrossRef] [Green Version]
- Dietz, T.H.; Byrne, R.A. Measurement of sulfate uptake and loss in the freshwater bivalve Dreissena polymorpha using a semi-microassay. Can. J. Zool 1999, 77, 331–336. [Google Scholar] [CrossRef]
- Potts, W.T.W. The inorganic composition of the blood of Mytilus edulis and Anodonta cygnea. J. Exp. Biol. 1954, 31, 376–385. [Google Scholar]
- Malley, D.F.; Huebner, J.D.; Donkersloot, K. Effects on ionic composition of blood and tissues ofAnodonta grandis grandis (Bivalvia) of an addition of aluminum and acid to a lake. Arch. Environ. Contam. Toxicol. 1988, 17, 479–491. [Google Scholar] [CrossRef]
- Andrews, P. Über den Blutchemismus des Flukrebses Orconectes limosus und seine Vernderung im Laufe des Jahres. J. Comp. Physiol. A 1967, 57, 7–43. [Google Scholar] [CrossRef]
- Wood, C.M.; McMahon, B. Mechanisms of acid-base and ionoregulation in white suckers (Catostomus commersoni) in natural soft water. J. Comp. Physiol. B 1984, 154, 35–46. [Google Scholar] [CrossRef]
- Nakada, T.; Zandi-Nejad, K.; Kurita, Y.; Kudo, H.; Broumand, V.; Kwon, C.Y.; Mercado, A.; Mount, D.B.; Hirose, S. Roles of Slc13a1 and Slc26a1 sulfate transporters of eel kidney in sulfate homeostasis and osmoregulation in freshwater. Am. J. Physiol. Integr. Comp. Physiol. 2005, 289, R575–R585. [Google Scholar] [CrossRef] [Green Version]
- Linker, P.H. Glycosaminoglycans in Anodonta californiensis, a Freshwater Mussel. Boil. Bull. 1993, 185, 263–276. [Google Scholar] [CrossRef]
- Luquet, G.; Fernández, M.S.; Badou, A.; Guichard, N.; Le Roy, N.; Corneillat, M.; Alcaraz, G.; Arias, J.L. Comparative Ultrastructure and Carbohydrate Composition of Gastroliths from Astacidae, Cambaridae and Parastacidae Freshwater Crayfish (Crustacea, Decapoda). Biomolecules 2012, 3, 18–38. [Google Scholar] [CrossRef]
- Yamada, S.; Sugahara, K.; Özbek, S. Evolution of glycosaminoglycans. Commun. Integr. Boil. 2011, 4, 150–158. [Google Scholar] [CrossRef]
- Cássaro, C.M.; Dietrich, C.P. Distribution of sulfated mucopolysaccharides in invertebrates. J. Boil. Chem. 1977, 252, 2254–2261. [Google Scholar]
- Hudson, B.H.; York, J.D. Roles for nucleotide phosphatases in sulfate assimilation and skeletal disease. Adv. Boil. Regul. 2012, 52, 229–238. [Google Scholar] [CrossRef] [Green Version]
- Klaassen, C.D.; Boles, J.W. The importance of 3′-phosphoadenosine 5‘-phosphosulfate (PAPS) in the regulation of sulfation. FASEB J. 1997, 11, 404–418. [Google Scholar] [CrossRef]
- Sun, M.; Leyh, T.S. Channeling in Sulfate Activating Complexes†. Biochemistry 2006, 45, 11304–11311. [Google Scholar] [CrossRef] [PubMed]
- Boom, J.V.D.; Heider, D.; Martin, S.R.; Pastore, A.; Mueller, J.W. 3’-Phosphoadenosine 5’-phosphosulfate (PAPS) synthases, naturally fragile enzymes specifically stabilized by nucleotide binding. J. Boil. Chem. 2012, 287, 17645–17655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bradley, T.J. Hyper-regulators: Life in fresh water. In Animal Osmoregulation; Oxford University Press (OUP): Oxford, UK, 2008; pp. 86–110. [Google Scholar]
- USEPA. Methods for Measuring the Acute Toxicity of Effluents and Receiving Waters to Freshwater and Marine Organisms; U.S. Environmental Protection Agency, Ed.; U.S. Environmental Protection Agency, Office of Water: Washington, DC, USA, 2002.
- Gordon, D.A.; Smith, M.E.; Wratschko, M.; Agard, D.; Holden, L.; Wilcox, S.; Lazorchak, J. A new approach for the laboratory culture of the fathead minnow, Pimephales promelas. Environ. Toxicol. Chem. 2013, 33, 126–133. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.E.; Lazorchak, J.; Herrin, L.E.; Brewer-Swartz, S.; Thoeny, W.T. A reformulated, reconstituted water for testing the freshwater amphipod, Hyalella azteca. Environ. Toxicol. Chem. 1997, 16, 1229–1233. [Google Scholar] [CrossRef]
- Wang, N.; Consbrock, R.A.; Ingersoll, C.G.; Hardesty, D.K.; Brumbaugh, W.G.; Hammer, E.; Bauer, C.R.; Mount, D.R. Acute and chronic toxicity of sodium sulfate to four freshwater organisms in water-only exposures. Environ. Toxicol. Chem. 2015, 35, 115–127. [Google Scholar] [CrossRef]
- Romano, N.; Zeng, C. Effects of potassium on nitrate mediated alterations of osmoregulation in marine crabs. Aquat. Toxicol. 2007, 85, 202–208. [Google Scholar] [CrossRef]
- Dietz, T.H.; Wilcox, S.J.; Silverman, H.; Byrne, R.A. Effects of hyperosmotic challenge on the freshwater bivalve Dreissena polymorpha: Importance of K+. Can. J. Zool 1997, 75, 697–705. [Google Scholar] [CrossRef]
- Henry, M.; Chester, D.; Mauck, W. Role of artificial burrows inHexageniatoxicity tests: Recommendations for protocol development. Environ. Toxicol. Chem. 1986, 5, 553–559. [Google Scholar] [CrossRef]
- Revesz, K.; Coplen, T.B. Determination of the δ34S of Total Sulfur in Solids: RSIL Lab Code 1800. In Techniques and Methods; US Geological Survey: Reston, VA, USA, 2006; pp. 1–31. [Google Scholar]
- USEPA. Methods for the Determination of Inorganic Substances in Environmental Samples; U.S. Environmental Protection Agency, Office of Research and Development: Cincinnati, OH, USA, 1993.
- Revesz, K.; Qi, H.; Coplen, T.B. Determination of the δ34S of sulfate in water; RSIL lab code 1951. In Techniques and Methods; US Geological Survey: Reston, VA, USA, 2006; pp. 1–31. [Google Scholar]
- Coplen, T.B. Guidelines and recommended terms for expression of stable-isotope-ratio and gas-ratio measurement results. Rapid Commun. Mass Spectrom. 2011, 25, 2538–2560. [Google Scholar] [CrossRef]
- Cobelli, C.; Toffolo, G.; Bier, D.M.; Nosadini, R. Models to interpret kinetic data in stable isotope tracer studies. Am. J. Physiol. Metab. 1987, 253, E551–E564. [Google Scholar] [CrossRef]
- Cobelli, C.; Toffolo, G.; Foster, D.M. Tracer-to-tracee ratio for analysis of stable isotope tracer data: Link with radioactive kinetic formalism. Am. J. Physiol. Metab. 1992, 262, E968–E975. [Google Scholar] [CrossRef] [PubMed]
- Wood, C.M. Acid-base and ionic exchanges at gills and kidney after exhaustive exercise in the rainbow trout. J. Exp. Biol. 1988, 136, 461–481. [Google Scholar]
- Wood, C.M.; Wheatly, M.G.; Hōbe, H. The mechanisms of acid-base and ionoregulation in the freshwater rainbow trout during environmental hyperoxia and subsequent normoxia. III. Branchial exchanges. Respir. Physiol. 1984, 55, 175–192. [Google Scholar] [CrossRef]
- Gonzalez, R.J.; Dunson, W.A. Adaptations of sodium balance to low pH in a sunfish (Enneacanthus obesus) from naturally acidic waters. J. Comp. Physiol. B 1987, 157, 555–566. [Google Scholar] [CrossRef]
- Robinson, R.B. Reference and intercomparison materials for stable isotopes of light elements. In Proceedings of the a Consultants Meeting, Vienna, Austria, 1–3 December 1993; International Atomic Energy Agency: Vienna, Austria, 1995; pp. 39–45. [Google Scholar]
- Stobbart, R.H. The effect of some anions and cations upon the fluxes and net uptake of chloride in the larva of Aëdes aegypti (L.), and the nature of the uptake mechanisms for sodium and chloride. J. Exp. Boil. 1967, 47, 35–57. [Google Scholar]
- Chen, Y.-Y.; Lu, F.-I.; Hwang, P.-P. Comparisons of calcium regulation in fish larvae. J. Exp. Zool 2003, 295, 127–135. [Google Scholar] [CrossRef]
- Wilcox, S.; Dietz, T. Potassium transport in the freshwater bivalve Dreissena polymorpha. J. Exp. Boil. 1995, 198, 861–868. [Google Scholar]
- Frain, W.J. The effect of external sodium and calcium concentrations on sodium fluxes by salt-depleted and non-depleted minnows, Phoxinus phoxinus (L.). J. Exp. Biol. 1987, 131, 417–425. [Google Scholar]
- Chasiotis, H.; Kolosov, D.; Kelly, S. Permeability properties of the teleost gill epithelium under ion-poor conditions. Am. J. Physiol. Integr. Comp. Physiol. 2012, 302, R727–R739. [Google Scholar] [CrossRef] [Green Version]
- Dietz, T.H.; Byrne, R.A.; Lynn, J.W.; Silverman, H. Paracellular solute uptake by the freshwater zebra mussel Dreissena polymorpha. Am. J. Physiol. Integr. Comp. Physiol. 1995, 269, R300–R307. [Google Scholar] [CrossRef]
- McDonald, D.G.; Rogano, M.S. Ion Regulation by the Rainbow Trout, Salmo gairdneri, in Ion-Poor Water. Physiol. Zool 1986, 59, 318–331. [Google Scholar] [CrossRef]
- Wood, C.M.; Gilmour, K.M.; Pärt, P. Passive and active transport properties of a gill model, the cultured branchial epithelium of the freshwater rainbow trout (Oncorhynchus mykiss). Comp. Biochem. Physiol. Part. A Mol. Integr. Physiol. 1998, 119, 87–96. [Google Scholar] [CrossRef]
- Takahashi, H.; Kopriva, S.; Giordano, M.; Saito, K.; Hell, R. Sulfur Assimilation in Photosynthetic Organisms: Molecular Functions and Regulations of Transporters and Assimilatory Enzymes. Annu. Rev. Plant. Boil. 2011, 62, 157–184. [Google Scholar] [CrossRef] [PubMed]
- McArthur, J.V.; Moorhead, K.K. Characterization of riparian species and stream detritus using multiple stable isotopes. Oecologia 1996, 107, 232–238. [Google Scholar] [CrossRef]
- Markovich, D.; Romano, A.; Storelli, C.; Verri, T. Functional and structural characterization of the zebrafish Na+-sulfate cotransporter 1 (NaS1) cDNA and gene (slc13a1). Physiol. Genom. 2008, 34, 256–264. [Google Scholar] [CrossRef]
- Katoh, F.; Tresguerres, M.; Lee, K.M.; Kaneko, T.; Aida, K.; Goss, G.G. Cloning of rainbow trout SLC26A1: Involvement in renal sulfate secretion. Am. J. Physiol. Integr. Comp. Physiol. 2006, 290, R1468–R1478. [Google Scholar] [CrossRef] [Green Version]
- Maddrell, S.H.P.; Phillips, J.E. Active transport of sulphate ions by the Malpighian tubules of larvae of the mosquito Aedes campestris. J. Exp. Biol. 1975, 62, 367–378. [Google Scholar]
- Maddrell, S.H.P.; Phillips, J.E. Induction of sulphate transport and hormonal control of fluid secretion by Malpighian Tubules of larvae of the mosquito Aedes taeniorhynchus. J. Exp. Biol. 1978, 72, 181–202. [Google Scholar]
- Gerencser, G.A.; Cattey, M.A.; Ahearn, G.A. Sulfate/oxalate exchange by lobster hepatopancreatic basolateral membrane vesicles. Am. J. Physiol. Integr. Comp. Physiol. 1995, 269, R572–R577. [Google Scholar] [CrossRef]
- Gerencser, G.A.; Ahearn, G.A.; Cattey, M.A. Antiport-driven sulfate secretion in an invertebrate epithelium. J. Exp. Zool 1996, 275, 269–276. [Google Scholar] [CrossRef]
- Cattey, M.A.; Gerencser, G.A.; Ahearn, G.A. Electrogenic H(+)-regulated sulfate-chloride exchange in lobster hepatopancreatic brush-border membrane vesicles. Am. J. Physiol. Integr. Comp. Physiol. 1992, 262, R255–R262. [Google Scholar] [CrossRef] [PubMed]
- Cattey, M.A.; Gerencser, G.A.; Ahearn, G.A. Electrogenic Coupling of Sulfate Secretion to Chloride Transport in Lobster Hepatopancreas. In Blood and Tissue Oxygen Carriers; Springer Science and Business Media LLC: Berlin, Germany, 1994; Volume 19, pp. 109–120. [Google Scholar]
- Horng, J.-L.; Hwang, P.-P.; Shih, T.-H.; Wen, Z.-H.; Lin, C.-S.; Lin, L.-Y. Chloride transport in mitochondrion-rich cells of euryhaline tilapia (Oreochromis mossambicus) larvae. Am. J. Physiol. Physiol. 2009, 297, C845–C854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choe, K.P.; O’Brien, S.; Evans, D.H.; Toop, T.; Edwards, S. Immunolocalization of Na+/K+-ATPase, carbonic anhydrase II, and vacuolar H+-ATPase in the gills of freshwater adult lampreys, Geotria australis. J. Exp. Zool 2004, 301, 654–665. [Google Scholar] [CrossRef] [PubMed]
- Filippov, V.; Aimanova, K.; Gill, S.S. Expression of an Aedes aegypti cation-chloride cotransporter and its Drosophila homologues. Insect Mol. Boil. 2003, 12, 319–331. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Wheatly, M.G. Characterization and expression of plasma membrane Ca2+ ATPase (PMCA3) in the crayfish Procambarus clarkii antennal gland during molting. J. Exp. Boil. 2004, 207, 2991–3002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ankley, G.T.; Bennett, R.; Erickson, R.J.; Hoff, D.J.; Hornung, M.W.; Johnson, R.D.; Mount, D.R.; Nichols, J.W.; Russom, C.L.; Schmieder, P.K.; et al. Adverse outcome pathways: A conceptual framework to support ecotoxicology research and risk assessment. Environ. Toxicol. Chem. 2010, 29, 730–741. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Mean SO42− influx (Jin) (A), efflux (Jout), and net flux (Jnet) (B) (μmol g−1 day−1) for each animal species—the fathead minnow, paper pondshell, red swamp crayfish, and two-lined mayfly—at the two target SO42− concentrations of 0.49 mmol L−1 and 175 mmol L−1. The error bars represent ±1 standard error. Because of some mortality occurring in the mayfly experiment, the numbers of replicates were 17 and 22 for the 0.49- and 1.25-mmol L−1 SO42− exposures, respectively. Moreover, an error in the initial water SO42− analysis in the mayfly experiment reduced the number of valid measurements for the initial 0.49 mmol L−1 water SO42−-concentration to 1, precluding a valid mean of Jout and Jnet for the 0.49 mmol L−1 SO42− exposure.
Figure 1.
Mean SO42− influx (Jin) (A), efflux (Jout), and net flux (Jnet) (B) (μmol g−1 day−1) for each animal species—the fathead minnow, paper pondshell, red swamp crayfish, and two-lined mayfly—at the two target SO42− concentrations of 0.49 mmol L−1 and 175 mmol L−1. The error bars represent ±1 standard error. Because of some mortality occurring in the mayfly experiment, the numbers of replicates were 17 and 22 for the 0.49- and 1.25-mmol L−1 SO42− exposures, respectively. Moreover, an error in the initial water SO42− analysis in the mayfly experiment reduced the number of valid measurements for the initial 0.49 mmol L−1 water SO42−-concentration to 1, precluding a valid mean of Jout and Jnet for the 0.49 mmol L−1 SO42− exposure.


{kind=link}
Table 1.
Concentrations of salts in the modified moderately hard reconstituted water (MMHRW) used in the experiment along with the resulting target mmol L−1 of the major ions, along with hardness and the molar [Na+]/[K+] ratios. Only the concentrations of Na+ and SO42− were increased in the MMHRW.
Table 1.
Concentrations of salts in the modified moderately hard reconstituted water (MMHRW) used in the experiment along with the resulting target mmol L−1 of the major ions, along with hardness and the molar [Na+]/[K+] ratios. Only the concentrations of Na+ and SO42− were increased in the MMHRW.
| Water | Salt | mg L−1 | Ions | mmol L−1 |
|---|---|---|---|---|
| Acclimation | CaCO3 | 71.0 | Na+ | 0.54 |
| NaHCO3 | 4.0 | K+ | 0.05 | |
| MgCl2·6H20 | 59.4 | Ca2+ | 0.71 | |
| KCl | 3.5 | Mg2+ | 0.29 | |
| Na2SO4 | 35.0 | HCO3− | 1.47 | |
| Cl− | 0.34 | |||
| SO42− | 0.25 | |||
| Hardness (mg/L, CaCO4) | 100.2 | |||
| Molar [Na+]/[K+] | 11.5 | |||
| 0.49 mmol L−1 SO42− | Na2SO4 | 70.0 | Na+ | 1.03 |
| SO42− | 0.49 | |||
| Hardness (mg/L, CaCO4) | 100.2 | |||
| Molar [Na+]/[K+] | 22.0 | |||
| 1.23 mmol L−1 SO42− | Na2SO4 | 175.0 | Na+ | 2.51 |
| SO42− | 1.23 | |||
| Hardness (mg L−1, CaCO4) | 100.2 | |||
| [Na+]/[K+] | 53.5 |
MMHRW = modified moderately hard reconstituted water.
Table 2.
Measured initial characteristics of SO42− in the test water for each species exposure. The column headings are the nominal concentrations of SO42−. The values are the sample mean ± 1 standard error. [SO42−] (mmol L−1) is the concentration of SO42−, and ẟ(34S/32S) (‰) is molar ratio of 34S/32S of the SO42−.
Table 2.
Measured initial characteristics of SO42− in the test water for each species exposure. The column headings are the nominal concentrations of SO42−. The values are the sample mean ± 1 standard error. [SO42−] (mmol L−1) is the concentration of SO42−, and ẟ(34S/32S) (‰) is molar ratio of 34S/32S of the SO42−.
| Variable | Species | 0.49 mmol L−1 | 1.23 mmol L−1 | ||
|---|---|---|---|---|---|
| n | Value | n | Value | ||
| [SO42−] (mmol L−1) | Fathead minnow | 5 | 0.52 ± 0.03 | 5 | 1.67 ± 0.03 |
| Paper pondshell | 5 | 0.35 ± 0.01 | 5 | 0.98 ± 0.06 | |
| Red swamp crayfish | 10 | 0.45 ± 0.01 | 10 | 1.15 ± 0.01 | |
| Two-lined mayfly | 1 | 0.48 | 3 | 1.19 ± 0.04 | |
| ẟ(34S/32S) (‰) | Fathead minnow | 5 | +1377.6 ± 1.3 | 5 | +1433.9 ± 0.8 |
| Paper pondshell | 5 | +1432.7 ± 34.1 | 5 | +1452.6 ± 0.5 | |
| Red swamp crayfish | 5 | +1475.6 ± 2.8 | 5 | +1461.3 ± 1.0 | |
| Two-lined mayfly | 5 | +1411.6 ± 3.0 | 5 | +1365.7 ± 4.4 | |
Table 3.
Proportion of variance in the J value means contributed by the variances of the mean values for component variables. The proportions of variance for other component variables were <0.001, or means were not used in the calculations. [S]int(0) is the mean calculated initial S concentration (μmol g−1) of each animal, and [SO42−]0 is the measured concentrations of SO42− in the water (μmol L−1) at the beginning of the exposure. Jin, Jout, and Jnet are the influx rates, efflux rates, and net flux of SO42− (μmol g−1 day−1), respectively, for each species.
Table 3.
Proportion of variance in the J value means contributed by the variances of the mean values for component variables. The proportions of variance for other component variables were <0.001, or means were not used in the calculations. [S]int(0) is the mean calculated initial S concentration (μmol g−1) of each animal, and [SO42−]0 is the measured concentrations of SO42− in the water (μmol L−1) at the beginning of the exposure. Jin, Jout, and Jnet are the influx rates, efflux rates, and net flux of SO42− (μmol g−1 day−1), respectively, for each species.
| Species | Target SO42− Concentration | J value | Variable | Proportion of Variance |
|---|---|---|---|---|
| Fathead minnow | 0.49 mmol L−1 | Jin | 0.603 | |
| Jout | 0.089 | |||
| 0.015 | ||||
| Jnet | 0.090 | |||
| 1.23 mmol L−1 | Jin | 0.584 | ||
| Jout | 0.583 | |||
| 0.006 | ||||
| Jnet | 0.058 | |||
| Paper pondshell | 0.49 mmol L−1 | Jin | 0.172 | |
| Jout | 0.046 | |||
| 0.021 | ||||
| Jnet | 0.047 | |||
| 1.23 mmol L−1 | Jin | 0.519 | ||
| Jout | 0.059 | |||
| 0.001 | ||||
| Jnet | 0.059 | |||
| Red swamp crayfish | 0.49 mmol L−1 | Jin | 0.582 | |
| 0.028 | ||||
| Jout | 0.012 | |||
| Jnet | 0.072 | |||
| 1.23 mmol L−1 | Jin | 0.747 | ||
| 0.002 | ||||
| Jout | 0.002 | |||
| Jnet | 0.005 | |||
| Two-lined mayfly | 0.49 mmol L−1 | Jin | 0.890 | |
| 1.23 mmol L−1 | Jin | 0.941 | ||
| Jout | 0.346 | |||
| 0.084 | ||||
| Jnet | 0.380 |
Table 4.
T-value (df and p) for the test of the H0 that each mean J = 0. mJin, mJout, and mJnet are the mean influx rate, mean efflux rate, and mean net flux of SO42−, respectively, for each species. Because of the multiple tests for each animal species, the Bonferroni adjustment of p is 0.0083. The t-values that indicate a mean J value statistically different from 0 are in bold.
Table 4.
T-value (df and p) for the test of the H0 that each mean J = 0. mJin, mJout, and mJnet are the mean influx rate, mean efflux rate, and mean net flux of SO42−, respectively, for each species. Because of the multiple tests for each animal species, the Bonferroni adjustment of p is 0.0083. The t-values that indicate a mean J value statistically different from 0 are in bold.
| Species | Target SO42− Concentration | H0: mJin = 0 t-Value (df, p) | H0: mJout = 0 t-Value (df, p) | H0: mJnet = 0 t-Value (df, p) |
|---|---|---|---|---|
| Fathead minnow | 0.49 mmol/L | 5.32 (24, <0.0001) | 27.88 (24, <0.0001) | 28.60 (24, <0.0001) |
| 1.23 mmol/L | 9.15 (24, <0.0001) | −49.38 (23, <0.0001) | −48.51 (23, <0.0001) | |
| Paper pondshell | 0.49 mmol/L | 2.98 (24, 0.0066) | −6.38 (24, <0.0001) | −6.04 (24, <0.0001) |
| 1.23 mmol/L | 5.91 (24, <0.0001) | −21.78 (24, <0.0001) | −21.74 (24, <0.0001) | |
| Red swamp crayfish | 0.49 mmol/L | 4.50 (24, 0.0001) | 19.26 (24, <0.0001) | 20.34 (24, <0.0001) |
| 1.23 mmol/L | 2.60 (24, 0.0155) | −4.80 (24, <0.0001) | −4.68 (24, <0.0001) | |
| Two-lined mayfly | 0.49 mmol/L | 1.95 (17, 0.0680) | ||
| 1.23 mmol/L | 1.97 (21, 0.0623) | −0.33 (2, 0.77) | −0.16 (2, 0.89) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Griffith, M.B.; Lazorchak, J.M.; Haring, H. Uptake of Sulfate from Ambient Water by Freshwater Animals. Water 2020, 12, 1496. https://doi.org/10.3390/w12051496
AMA Style
Griffith MB, Lazorchak JM, Haring H. Uptake of Sulfate from Ambient Water by Freshwater Animals. Water. 2020; 12(5):1496. https://doi.org/10.3390/w12051496
Chicago/Turabian StyleGriffith, Michael B., James M. Lazorchak, and Herman Haring. 2020. "Uptake of Sulfate from Ambient Water by Freshwater Animals" Water 12, no. 5: 1496. https://doi.org/10.3390/w12051496
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.