Parameterization and Application of Stanghellini Model for Estimating Greenhouse Cucumber Transpiration

 , ,
, ,
Abstract
:Novelty Statement
Abstract
1. Introduction
2. Materials and Methods
2.1. Greenhouse Description and Site
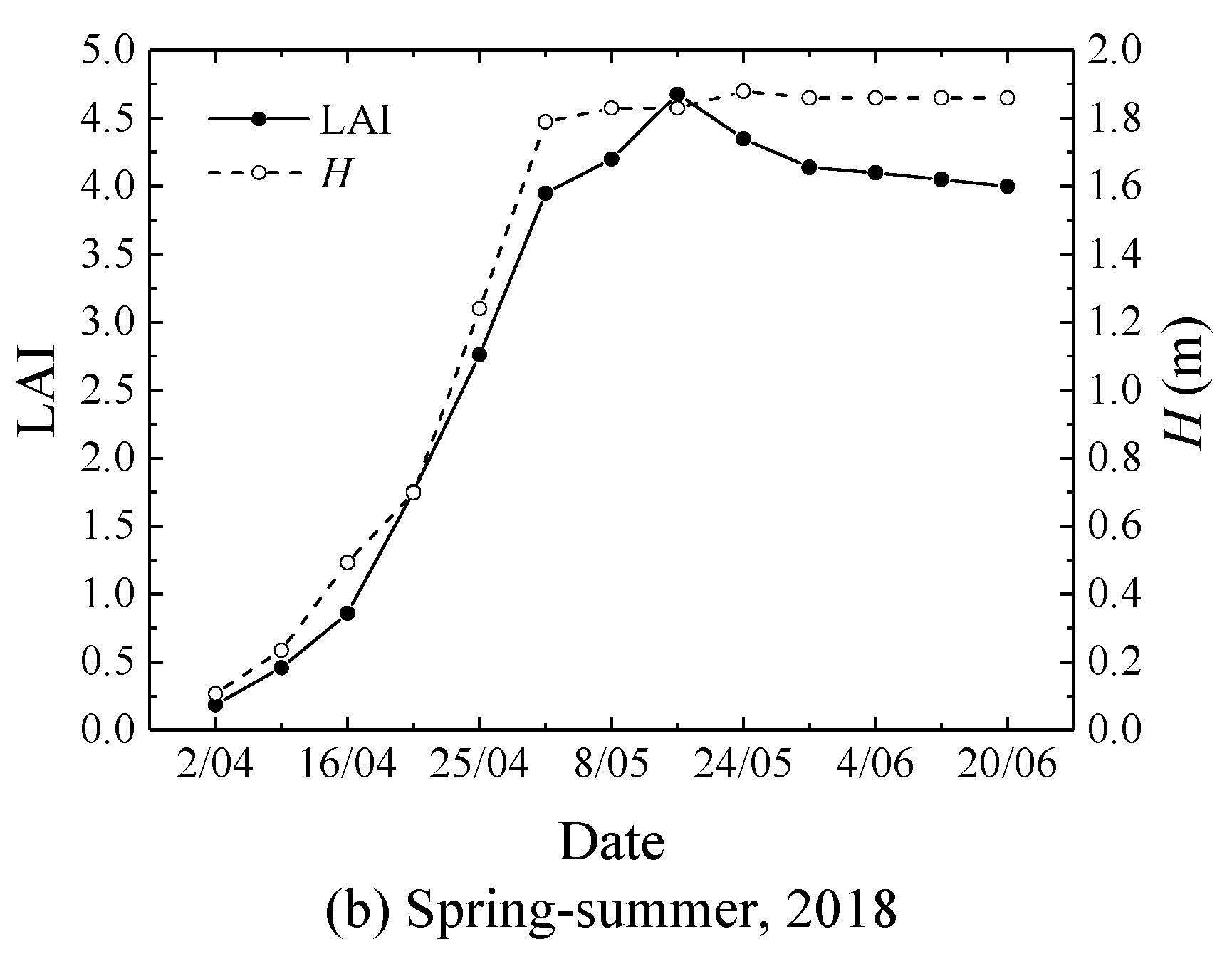
2.2. Crop
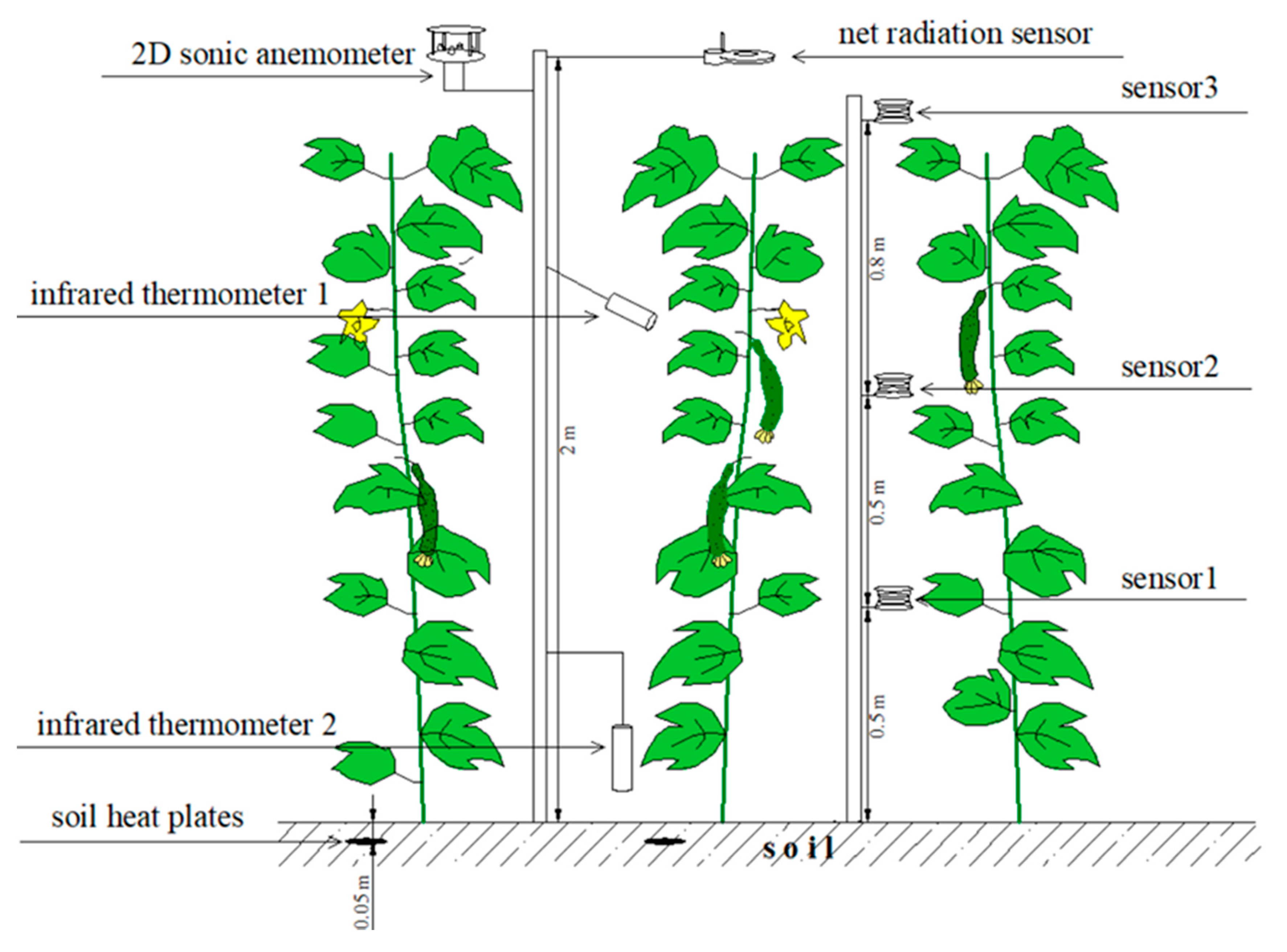
2.3. Instrumentation
2.4. Theoretical Model
2.4.1. Stanghellini Model (SM)
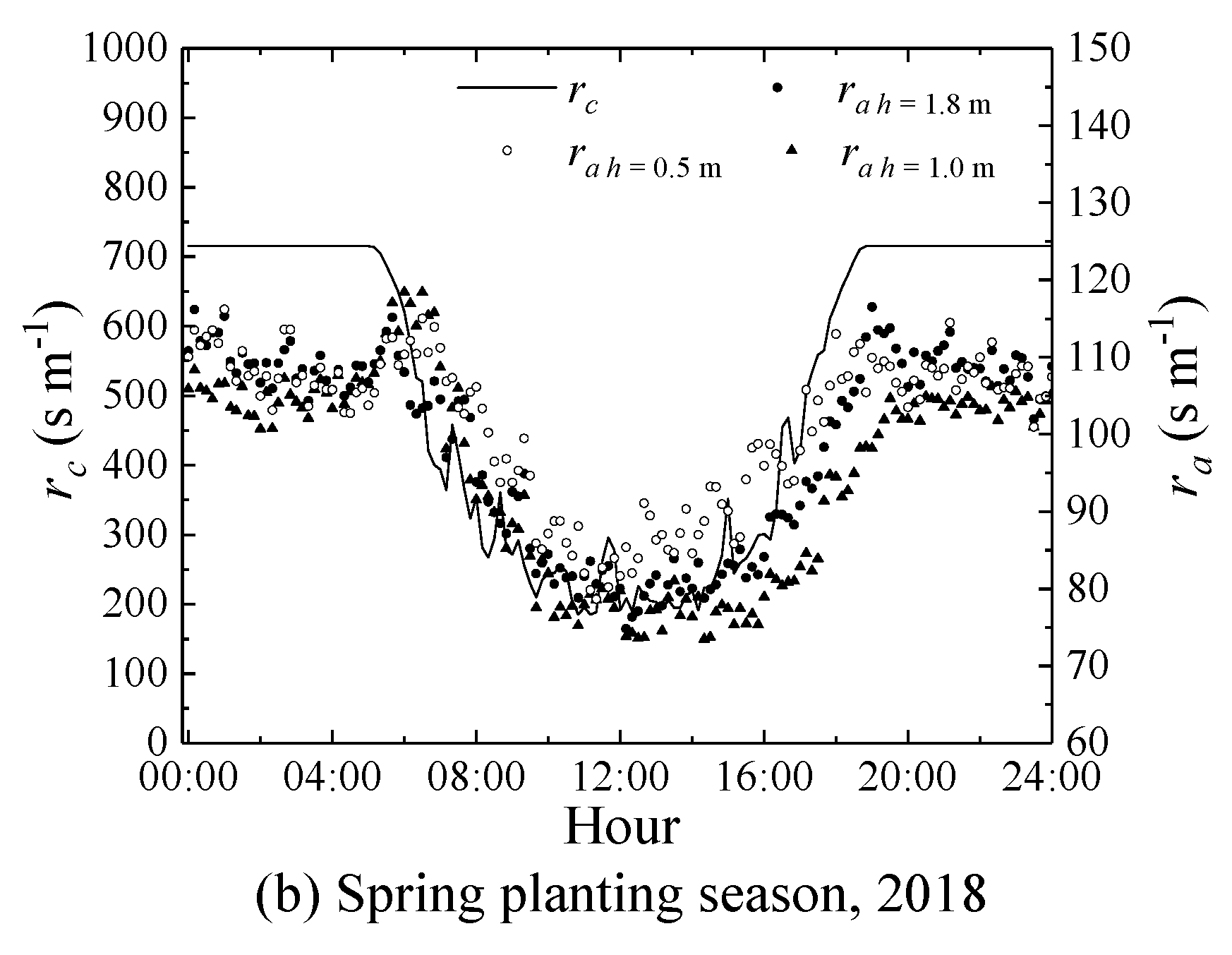
2.4.2. Aerodynamic Resistance and Canopy Resistance
2.4.3. Statistical Analysis
3. Results and Discussions
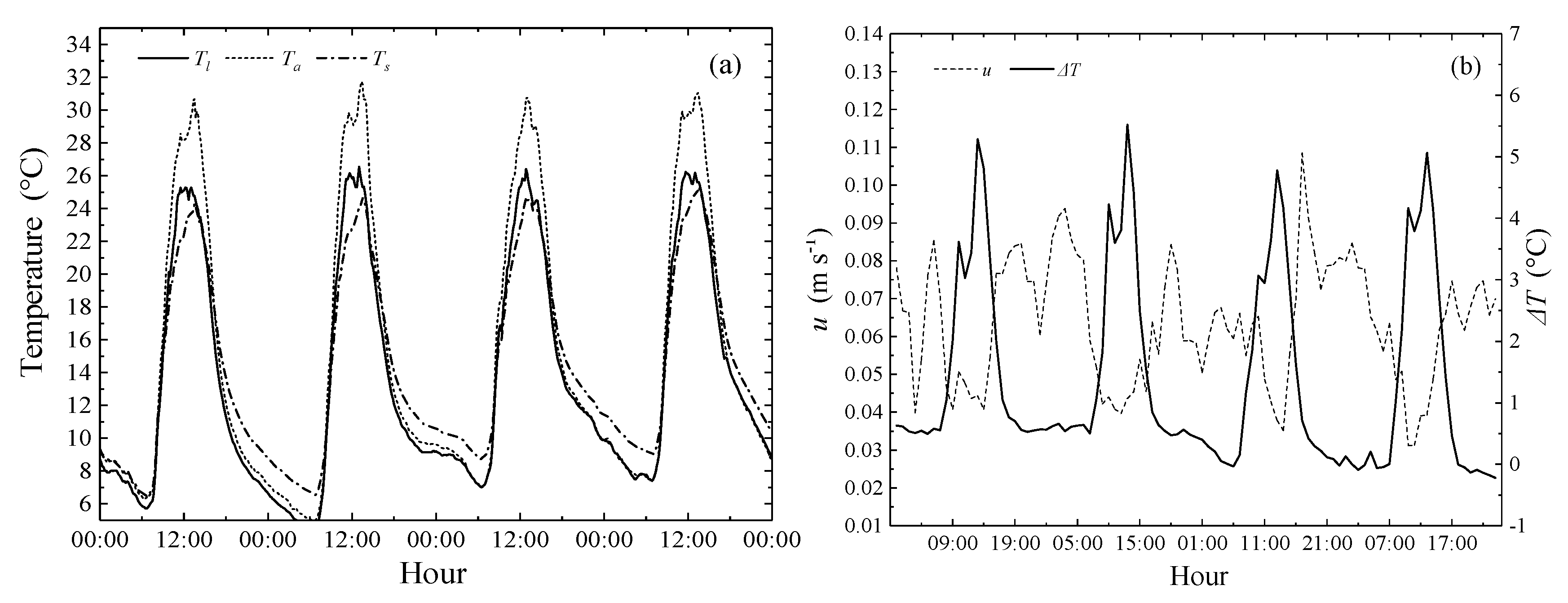
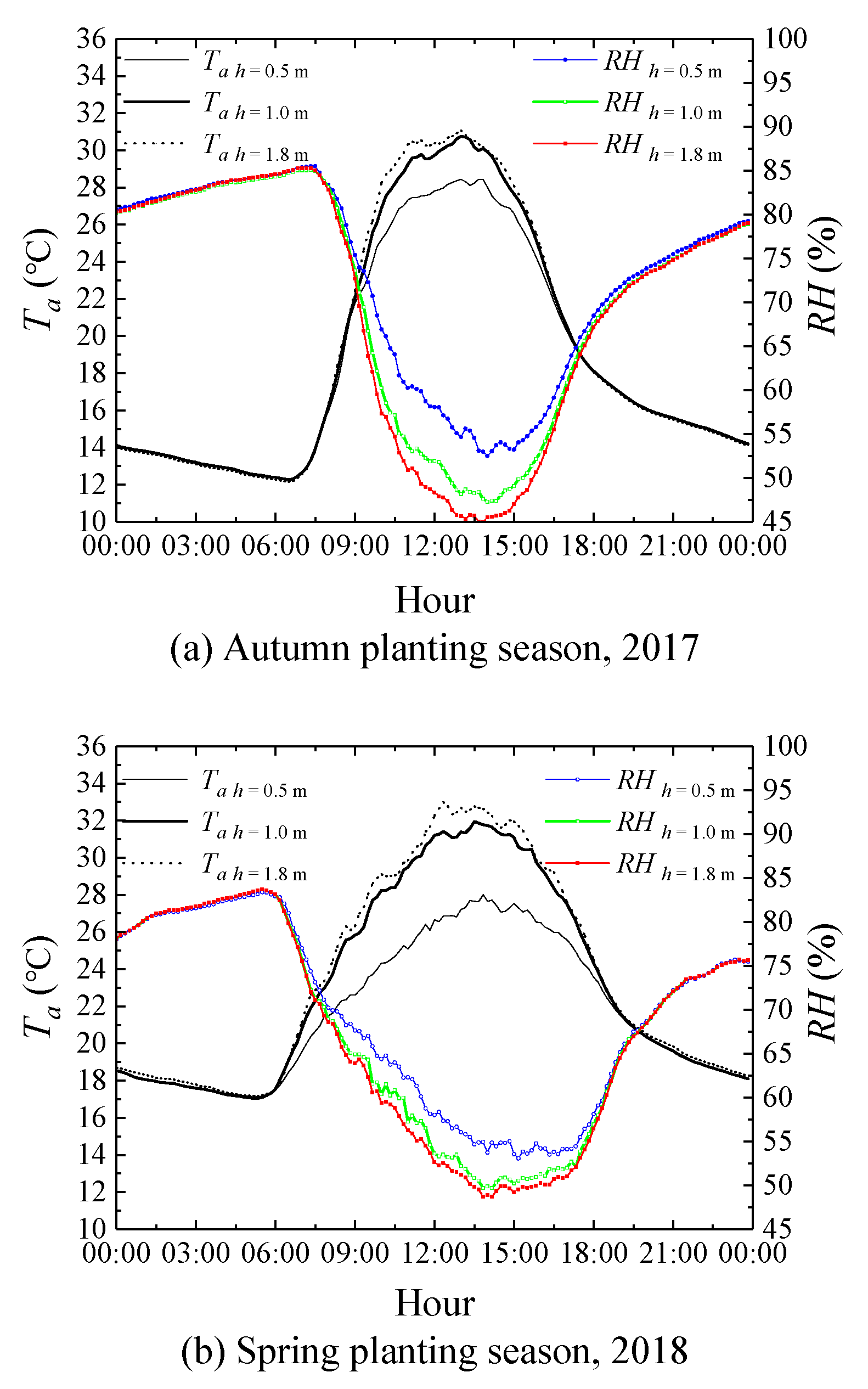
3.1. Meteorological Characterization in Greenhouse
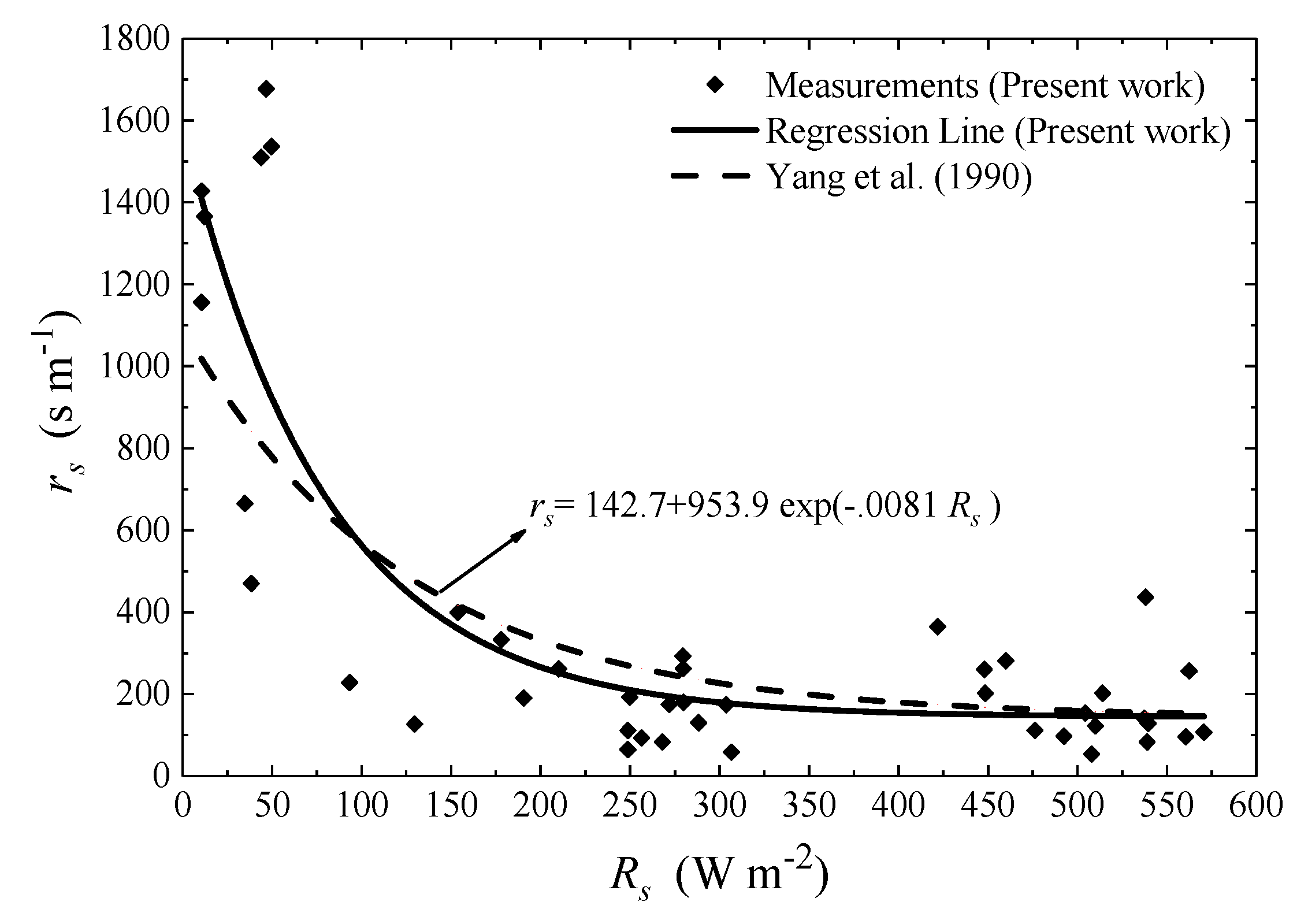
3.2. Parametrization of the Stanghellini Model (SM)
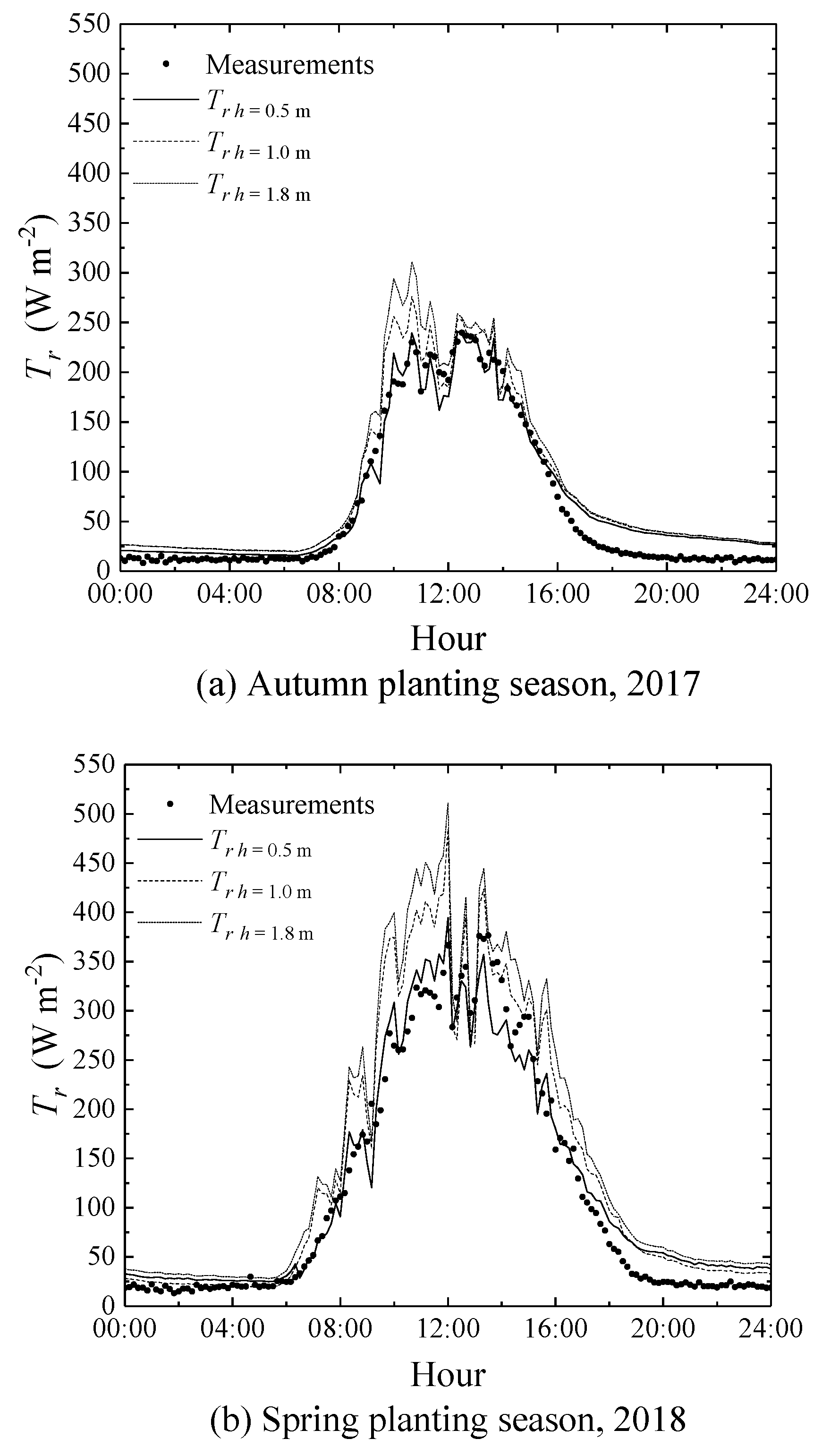
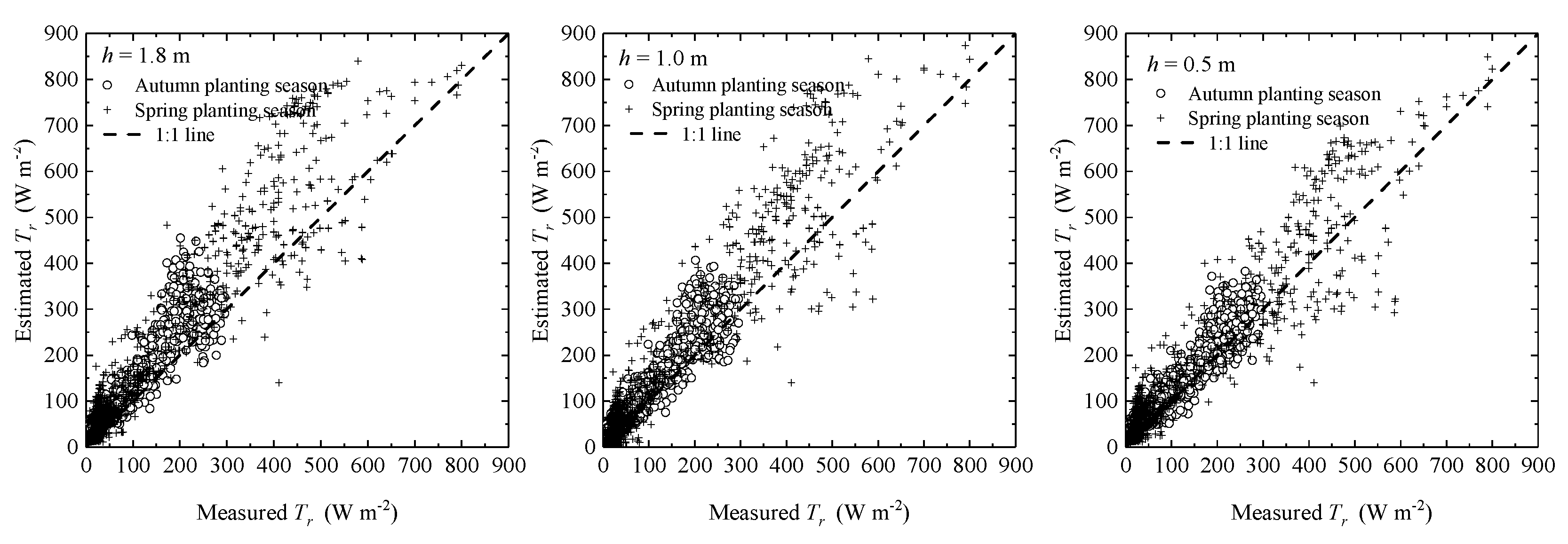
3.3. Effects of Micrometeorological Data Observation Heights on the Performance of the SM
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Luo, Y.F.; Traore, S.; Lyu, X.; Wang, W.G.; Wang, Y.; Xie, Y.Y.; Jiao, X.; Fipps, G. Medium Range Daily Reference Evapotranspiration Forecasting by Using ANN and Public Weather Forecasts. Water Resour. Manag. 2015, 29, 3863–3876. [Google Scholar] [CrossRef]
- Han, W.T.; Sun, Y.; Xu, T.F.; Chen, X.W.; Su, K.O. Detecting maize leaf water status by using digital rgb images. Int. J. Agric. Biol. Eng. 2014, 7, 45–53. [Google Scholar]
- Xu, D.; Li, Y.N.; Gong, S.H.; Zhang, B.Z. Waterlogging and saline-alkaline management for development of sustainably irrigated agriculture. J. Drain. Irrig. Mach. Eng. 2019, 37, 63–72, (In Chinese with English abstract). [Google Scholar]
- De, J.S.; Guo, P.; Zhang, C.L.; Yue, Q.; Shan, B.Y. Optimal allocation of irrigation water resources based on meteorological factor under uncertainty. J. Drain. Irrig. Mach. Eng. 2019, 37, 540–544, (In Chinese with English abstract). [Google Scholar]
- Allen, R.G.; Pereira, L.S.; Raes, D.; Smith, M. Crop Evapotranspiration—Guidelines for Computing Crop Water Requirements; FAO Irrigation and Drainage Paper 56; FAO: Rome, Italy, 1998; p. 300. [Google Scholar]
- Penman, H.L. Natural evaporation from open water, bare soil and grass. Proc. R. Soc. London A Math. Phys. Eng. Sci. 1948, 1032, 120–145. [Google Scholar]
- Monteith, J.L. Evaporation and environment. In Symposia of the Society for Experimental Biology; University Press: Cambridge, UK, 1965; pp. 206–234. [Google Scholar]
- Stanghellini, C. Transpiration of Greenhouse Crops: An Aid to Climate Management. Ph.D. Thesis, Agricultural University of Wageningen, Wageningen, The Netherlands, 1987; 150p. [Google Scholar]
- Villarreal-Guerrero, F.; Kacira, M.; Fitz-Rodríguez, E.; Kubota, C.; Giacomelli, G.A.; Linker, R.; Arbel, A. Comparison of three evapotranspiration models for a greenhouse cooling strategy with natural ventilation and variable high pressure fogging. Sci. Hortic. 2012, 134, 210–221. [Google Scholar] [CrossRef]
- Jolliet, O.; Bailey, B.J. The effect of climate on tomato transpiration in greenhouses: Measurements and models comparison. Agric. For. Meteorol. 1992, 58, 43–62. [Google Scholar] [CrossRef]
- Prenger, J.J.; Fynn, R.P.; Hansen, R.C. A comparison of four evapotranspiration models in a greenhouse environment. Trans. ASAE 2002, 45, 1779–1788. [Google Scholar] [CrossRef]
- Lopez-Cruz, I.L.; Olivera-Lopez, M.; Herrera-Ruiz, G. Simulation of greenhouse tomato crop transpiration by two theoretical models. Acta Hortic. 2008, 797, 145–150. [Google Scholar] [CrossRef]
- Pamungkas, A.P.; Hatou, K.; Morimoto, T. Evapotranspiration model analysis of crop water use in plant factory system. Environ. Control Biol. 2014, 52, 183–188. [Google Scholar] [CrossRef] [Green Version]
- Demrati, H.; Boulard, T.; Fatnassi, H.; Bekkaoui, A.; Majdoubi, H.; Elattir, H.; Bouirden, L. Microclimate and transpiration of a greenhouse banana crop. Biosyst. Eng. 2007, 98, 66–78. [Google Scholar] [CrossRef]
- Morille, B.; Migeon, C.; Bournet, P.E. Is the Penman-Monteith model adapted to predict crop transpiration under greenhouse conditions? Application to a New Guinea Impatiens crop. Sci. Hortic. 2013, 152, 80–91. [Google Scholar] [CrossRef]
- Yan, H.F.; Zhang, C.; Oue, H.; Peng, G.J.; Darko, R.O. Determination of crop and soil evaporation coefficients for estimating evapotranspiration in a paddy field. Int. J. Agric. Biol. Eng. 2017, 10, 130–139. [Google Scholar]
- Katerji, N.; Rana, G.; Fahed, S. Parameterizing canopy resistance using mechanistic and semi-empirical estimates of hourly evapotranspiration: Critical evaluation for irrigated crops in the Mediterranean. Hydrol. Process. 2011, 25, 117–129. [Google Scholar] [CrossRef]
- Yan, H.F.; Zhang, C.; Peng, G.J.; Darko, R.; Cai, B. Modelling canopy resistance for estimating latent heat flux at a tea field in south China. Exp. Agric. 2018, 54, 563–576. [Google Scholar] [CrossRef]
- Yang, X.S.; Short, T.H.; Fox, R.D.; Bauerle, W.L. Transpiration, leaf temperature and stomatal resistance of greenhouse cucumber crop. Agric. For. Meteorol. 1990, 51, 197–209. [Google Scholar] [CrossRef]
- Qiu, R.J.; Kang, S.Z.; Du, T.S.; Tong, L.; Hao, X.M.; Chen, R.Q.; Chen, J.L.; Li, F.S. Effect of convection on the Penman-Monteith model estimates of transpiration of hot pepper grown in solar greenhouse. Sci. Hortic. 2013, 160, 163–171. [Google Scholar] [CrossRef]
- Gong, X.W.; Liu, H.; Sun, J.S.; Gao, Y.; Zhang, X.X.; Shiva, K.J.H.A.; Zhang, H.; Ma, X.J.; Wang, W.N. A proposed surface resistance model for the Penman-Monteith formula to estimate evapotranspiration in a solar greenhouse. J. Arid Land 2017, 9, 530–546. [Google Scholar] [CrossRef]
- Jarvis, P.G. The interpretation of the variations in leaf water potential and stomatal conductance found in canopies in the field. Philos. Trans. R. Soc. B Biol. Sci. 1976, 273, 593–610. [Google Scholar]
- Yang, X.S. Greenhouse micrometeorology and estimation of heat and water vapor fluxes. J. Agric. Eng. Res. 1995, 61, 227–237. [Google Scholar] [CrossRef]
- Kittas, C.; Katsoulas, N.; Baille, A. Influence of misting on the diurnal hysteresis of canopy transpiration rate and conductance in a rose greenhouse. Acta Hortic. 2000, 534, 155–161. [Google Scholar] [CrossRef]
- Yan, H.F.; Zhang, C.; Gerrits, M.C.; Acquah, S.J.; Zhang, H.N.; Wu, H.M.; Zhao, B.S.; Huang, S.; Fu, H.W. Parametrization of aerodynamic and canopy resistances for modeling evapotranspiration of greenhouse cucumber. Agric. For. Meteorol. 2018, 262, 370–378. [Google Scholar] [CrossRef]
- Huang, S.; Yan, H.F.; Zhang, C.; Wang, G.Q.; Acquah, S.J.; Yu, J.J.; Li, L.L.; Ma, J.M.; Opoku Darko, R. Modeling evapotranspiration for cucumber plants based on the Shuttleworth-Wallace model in a Venlo-type greenhouse. Agric. Water Manag. 2020, 228, 105861. [Google Scholar] [CrossRef]
- Yan, H.F.; Acquah, S.J.; Zhang, C.; Wang, G.Q.; Huang, S.; Zhang, H.N.; Zhao, B.S.; Wu, H.M. Energy partitioning of greenhouse cucumber based on the application of Penman-Monteith and Bulk Transfer models. Agric. Water Manag. 2019, 217, 201–211. [Google Scholar] [CrossRef]
- Liu, H. Water Requirement and Optimal Irrigation Index for Effective Water Use and High-Quality of Tomato in Greenhouse. Ph.D. Thesis, Chinese Academy of Agricultural Sciences, Beijing, China, 2010. (In Chinese with English abstract). [Google Scholar]
- Montero, J.I.; Anton, A.; Munoz, P.; Lorenzo, P. Transpiration from geranium grown under high temperatures and low humidities in greenhouses. Agric. For. Meteorol. 2001, 107, 323–332. [Google Scholar] [CrossRef]
- Pirkner, M.; Dicken, U.; Tanny, J. Penman-Monteith approaches for estimating crop evapotranspiration in screenhouses—A case study with table-grape. Int. J. Biometeorol. 2014, 58, 725–737. [Google Scholar] [CrossRef]
- Papadakis, G.; Frangoudakis, A.; Kyritsis, S. Experimental investigation and modelling of heat and mass transfer between a tomato crop and the greenhouse environment. J. Agric. Eng. Res. 1994, 57, 217–227. [Google Scholar] [CrossRef]
- Acquah, S.J.; Yan, H.F.; Zhang, C.; Wang, G.Q.; Zhao, B.S.; Wu, H.M.; Zhang, H.N. Application and evaluation of Stanghellini model in the determination of crop evapotranspiration in a naturally ventilated greenhouse. Int. J. Agric. Biol. Eng. 2018, 11, 95–103. [Google Scholar]
- Wang, H.H.; Zhou, G.M.; Li, X.Y. Heat Transfer Theory; Chongqing University Press: Chongqing, China, 2006; 185p. [Google Scholar]
- Zhang, L.; Lemeur, R. Effect of aerodynamic resistance on energy balance and Penman—Monteith estimates of evapotranspiration in solar greenhouse conditions. Agric. For. Meteorol. 1992, 58, 209–228. [Google Scholar] [CrossRef]
- Allen, R.G.; Pereira, L.S.; Howell, T.A.; Jensen, M.E. Evapotranspiration information reporting: I. Factors governing measurement accuracy. Agric. Water Manag. 2011, 98, 899–920. [Google Scholar] [CrossRef] [Green Version]
- Choudhury, B.U.; Singh, A.K.; Pradhan, S. Estimation of crop coefficients of dry-seeded irrigated rice—Wheat rotation on raised beds by field water balance method in the indo-gangetic plains, India. Agric. Water Manag. 2013, 123, 20–31. [Google Scholar] [CrossRef]
- Tian, F.; Hou, M.; Qiu, Y.; Zhang, T.; Yuan, Y. Salinity stress effects on transpiration and plant growth under different salinity soil levels based on thermal infrared remote (TIR) technique. Geoderma 2020, 357, 113961. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Measured Heights | estimated | measured | a | R2 | RMSE | EF |
|---|---|---|---|---|---|---|
| h = 1.8 m | 123.08 | 91.40 | 1.29 | 0.92 | 42.86 | 83.29% |
| h = 1.0 m | 112.85 | 91.40 | 1.20 | 0.94 | 31.71 | 90.01% |
| h = 0.5 m | 108.11 | 91.40 | 0.97 | 0.91 | 26.19 | 93.19% |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yan, H.; Huang, S.; Zhang, C.; Gerrits, M.C.; Wang, G.; Zhang, J.; Zhao, B.; Acquah, S.J.; Wu, H.; Fu, H. Parameterization and Application of Stanghellini Model for Estimating Greenhouse Cucumber Transpiration. Water 2020, 12, 517. https://doi.org/10.3390/w12020517
Yan H, Huang S, Zhang C, Gerrits MC, Wang G, Zhang J, Zhao B, Acquah SJ, Wu H, Fu H. Parameterization and Application of Stanghellini Model for Estimating Greenhouse Cucumber Transpiration. Water. 2020; 12(2):517. https://doi.org/10.3390/w12020517
Chicago/Turabian StyleYan, Haofang, Song Huang, Chuan Zhang, Miriam Coenders Gerrits, Guoqing Wang, Jianyun Zhang, Baoshan Zhao, Samuel Joe Acquah, Haimei Wu, and Hanwen Fu. 2020. "Parameterization and Application of Stanghellini Model for Estimating Greenhouse Cucumber Transpiration" Water 12, no. 2: 517. https://doi.org/10.3390/w12020517