A Cone Fishway Facilitates Lateral Migrations of Tropical River-Floodplain Fish Communities

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
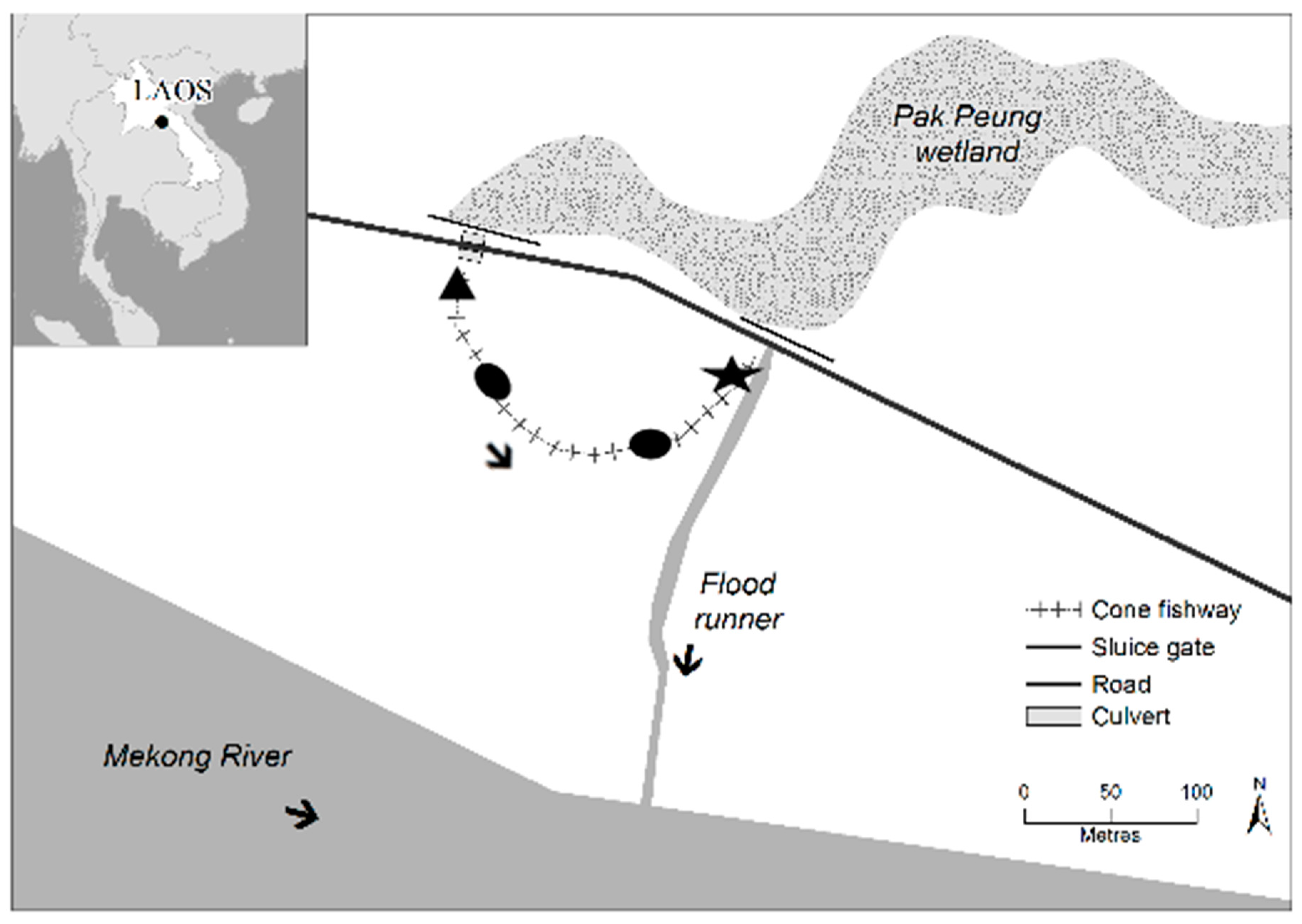
2.1. Study Site
2.2. Experimental Design
2.3. Data Analysis
3. Results
3.1. Overall Results
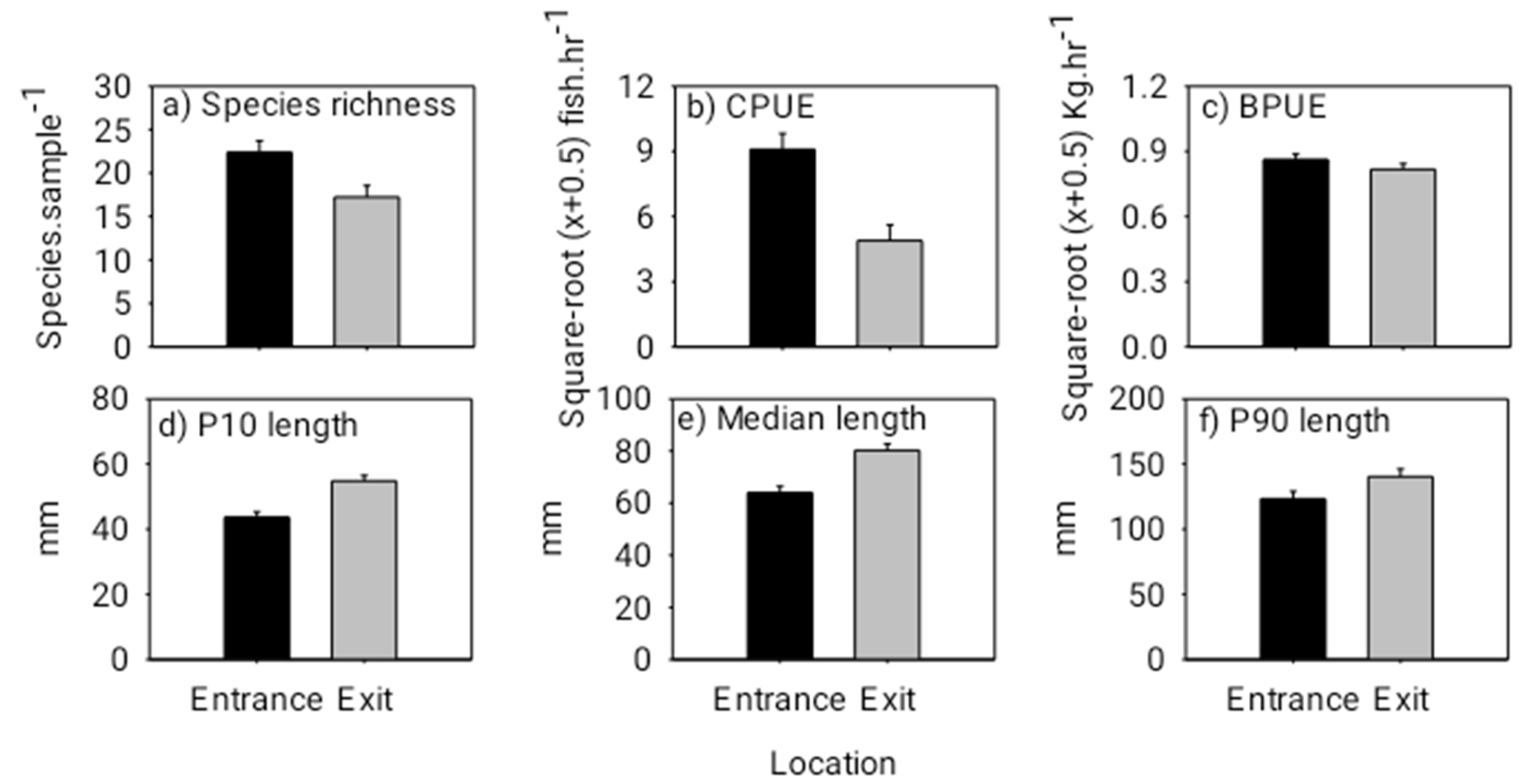
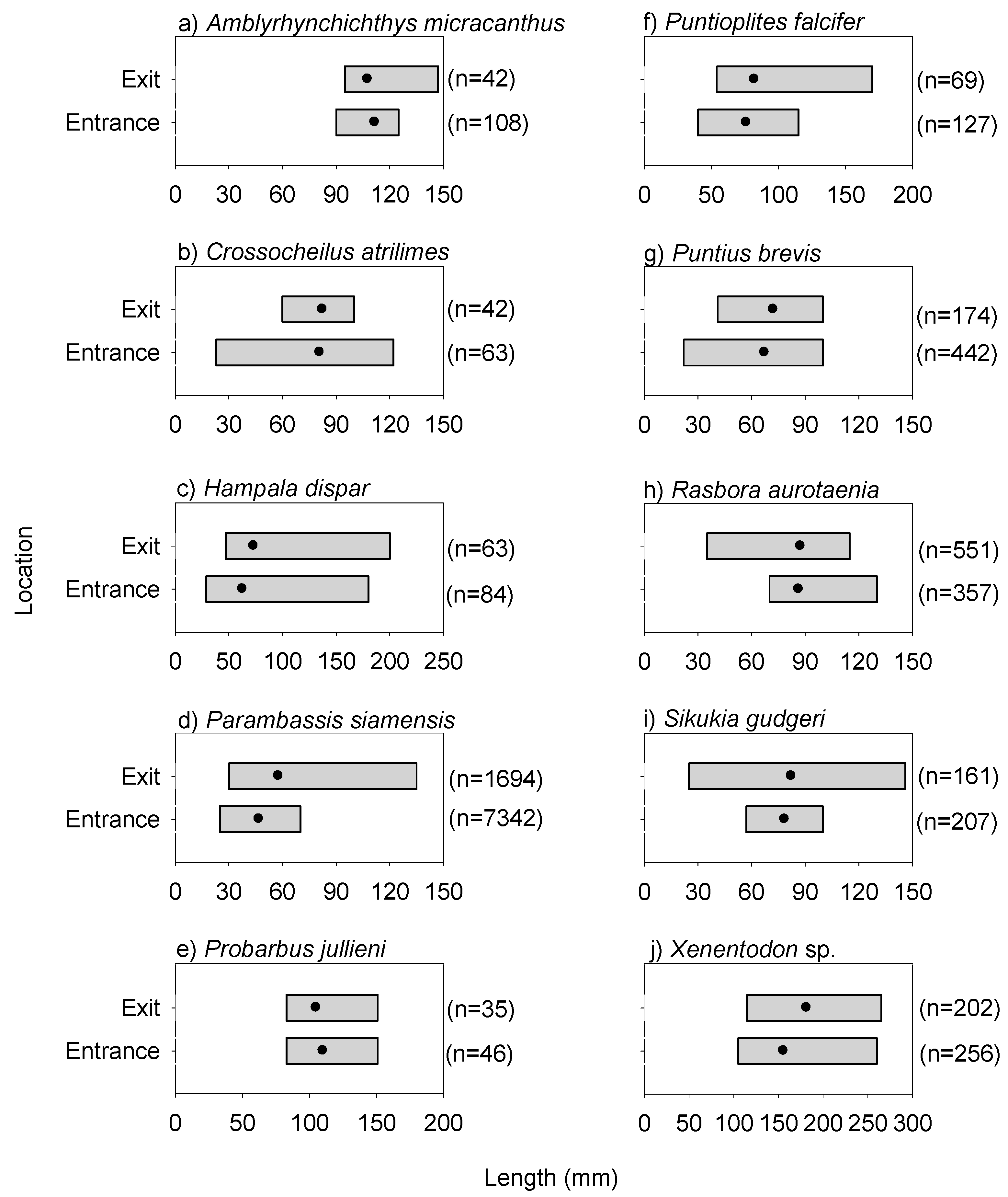
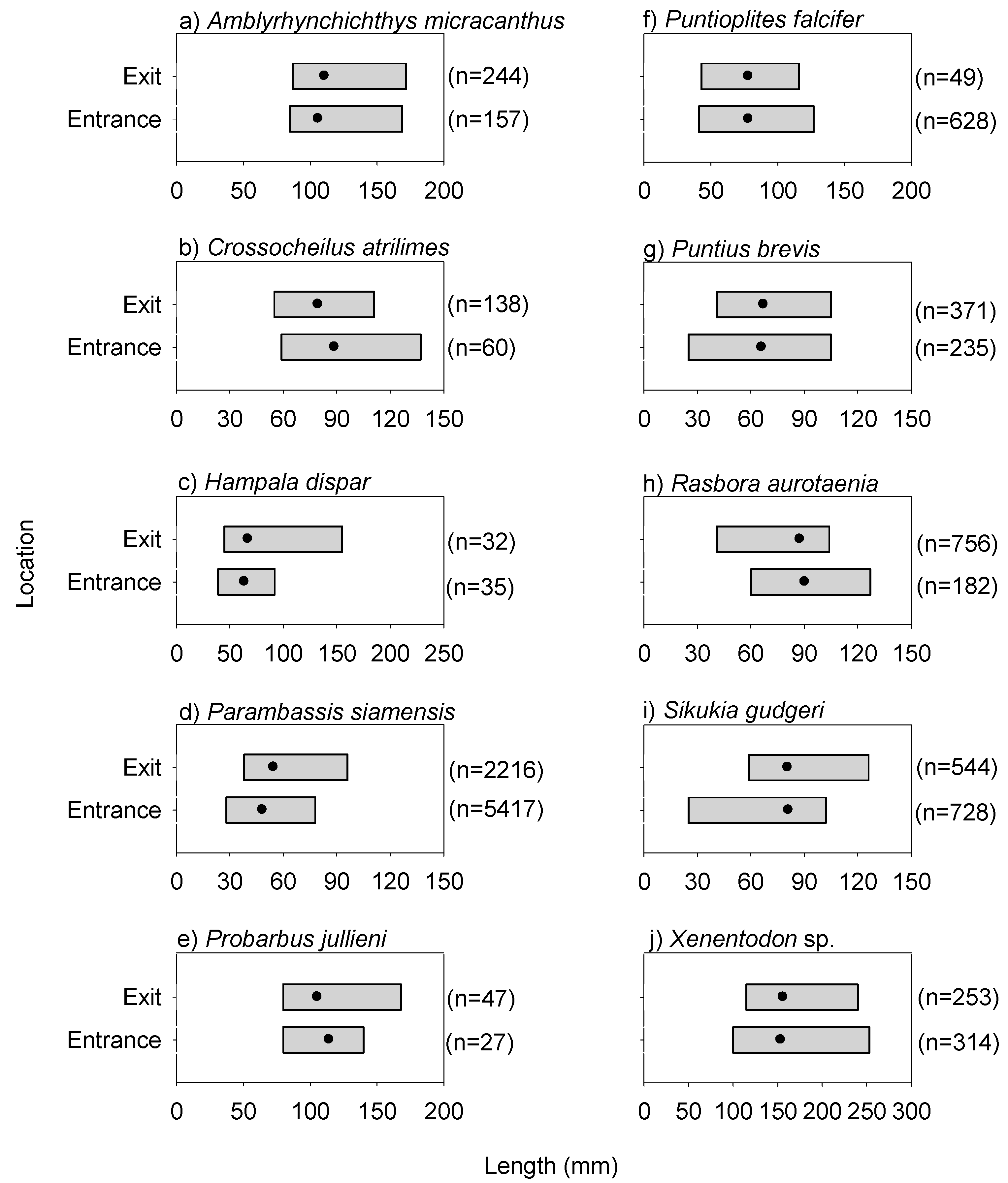
3.2. Fishway Entrance and Exit Patterns from the Day Experiment
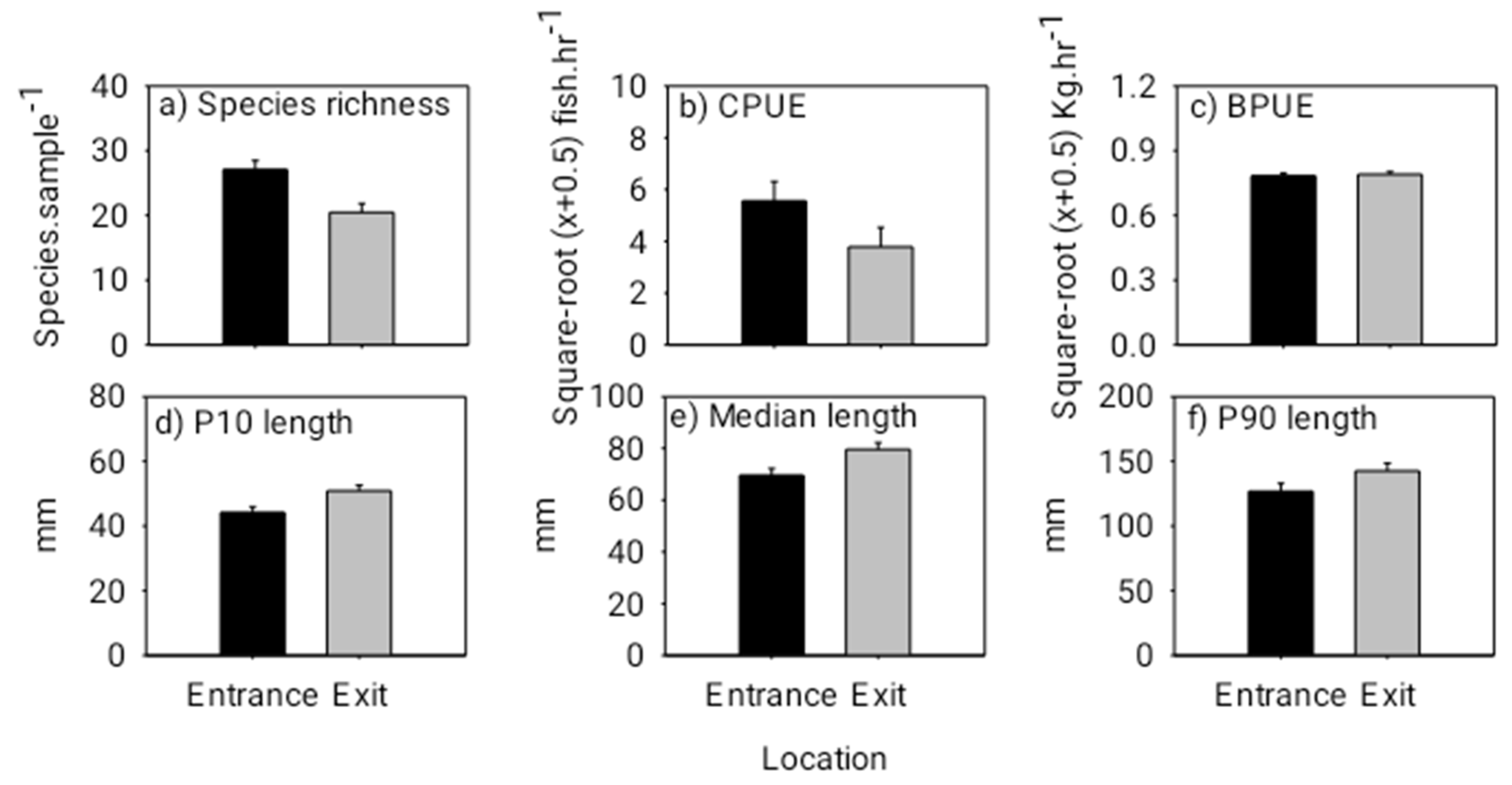
3.3. Fishway Entrance and Exit Patterns from the Dusk/Night Experiment
3.4. Fish Biomass Transfer facilitated by the Cone Fishway
4. Discussion
4.1. Effectiveness of the Cone Fishway Design
4.2. Diurnal Patterns
4.3. Management Implications and Areas for Future Research
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Total N | Day Experiment | Dusk/Night Experiment | ||
|---|---|---|---|---|---|
| Entrance | Exit | Entrance | Exit | ||
| Acanthopsis spp. | 7 | 140–155 | 149–149 | 121–152 | 138–138 |
| Akysis ephippifer | 8 | 27–42 | |||
| Amblyrhynchichthys micracanthus | 551 | 90–125 | 95–147 | 85–169 | 87–172 |
| Amblyrhynchichthys truncatus | 2 | 107–122 | |||
| Anabas testudineus | 203 | 40–95 | 155–162 | 40–63 | 119–119 |
| Badis ruber | 5 | 39–45 | 35–41 | ||
| Barbonymus altus | 199 | 25–102 | 40–86 | 35–95 | 40–103 |
| Channa gachua | 1 | 174–174 | |||
| Channa striata | 57 | 25–285 | 91–91 | 69–250 | |
| Chela laubuca | 1 | 80–80 | |||
| Chitala blanci | 1 | 100–100 | |||
| Chitala ornata | 23 | 83–140 | 110–157 | ||
| Cirrhinus cirrhosus | 12 | 80–111 | 71–71 | 55–120 | 50–50 |
| Clarias macrocephalus | 3 | 39–106 | 112–112 | ||
| Clarias sp. (cf. batrachus) | 2 | 134–134 | 105–105 | ||
| Clupeichthys aesarnensis | 3456 | 34–64 | 52–65 | 35–69 | 40–54 |
| Crossocheilus atrilimes | 303 | 23–122 | 60–100 | 59–137 | 55–111 |
| Cyclocheilichthys apogon | 3 | 69-69 | 80–105 | ||
| Cyclocheilichthys armatus | 22 | 70–115 | 106–111 | 67–100 | 70–111 |
| Cyclocheilichthys enoplos | 28 | 81–144 | 92–154 | 98–111 | 95–117 |
| Cyclocheilichthys lagleri | 34 | 77–115 | 90–130 | 42–114 | 65–120 |
| Cyprinus carpio | 9 | 80–119 | 100–100 | 116–139 | 98–98 |
| Esomus metallicus | 409 | 45–82 | 31–85 | 42–82 | |
| Hampala dispar | 214 | 29–180 | 47–200 | 39–92 | 45–155 |
| Hampala macrolepidota | 26 | 43–73 | 54–168 | 65–83 | 57–82 |
| Hemibagrus nemurus | 8 | 195–220 | 165–230 | ||
| Hemibagrus spp. | 20 | 56–155 | 194–240 | ||
| Henicorhynchus lobatus | 96 | 60–133 | 65–170 | 54–135 | 55–145 |
| Henicorhynchus ornatipinnis | 30 | 80–119 | 78–102 | 71–98 | 65–96 |
| Henicorhynchus siamensis | 449 | 31–95 | 40–185 | 49–112 | 55–145 |
| Homaloptera smithi | 193 | 36–36 | 33–33 | 29–40 | 30–41 |
| Hypophthalmichthys nobilis | 1 | 92–92 | |||
| Hypsibarbus lagleri | 170 | 36–111 | 43–125 | 37–105 | 46–163 |
| Hypsibarbus malcolmi | 63 | 80–128 | 77–141 | 75–135 | 79–127 |
| Hypsibarbus wetmorei | 14 | 67–90 | 90–130 | 105–150 | |
| Kryptopterus bicirrhis | 1 | 152–152 | |||
| Kryptopterus cryptopterus | 2 | 145–145 | 175–175 | ||
| Labiobarbus leptocheilus | 709 | 90–162 | 83–195 | 69–160 | 51–168 |
| Labiobarbus siamensis | 8 | 99–115 | 95–130 | ||
| Laides longibarbis | 1 | 90–90 | |||
| Macrognathus semiocellatus | 63 | 145–221 | 145–175 | 62–225 | 130–155 |
| Macrognathus siamensis | 57 | 80–231 | 170–202 | 109–198 | 152–190 |
| Mastacembelus armatus | 12 | 135–215 | 134–205 | 230–230 | |
| Mastacembelus favus | 6 | 120-315 | 140-300 | ||
| Monopterus albus | 3 | 63–109 | 220–220 | ||
| Mystacoleucus ectypus | 7 | 45–65 | 68–68 | 57–57 | |
| Mystacoleucus marginatus | 29 | 43–70 | 52–70 | 45–69 | 57–62 |
| Mystus albolineatus | 6 | 187–187 | 161–250 | 125–216 | |
| Mystus atrifasciatus | 40 | 52–85 | 63–89 | 45–78 | 40–180 |
| Mystus mysticetus | 2 | 115–115 | 122–122 | ||
| Mystus singaringan | 2 | 175-200 | |||
| Nandus oxyrhynchus | 16 | 60–92 | 80–80 | 50–78 | 63–82 |
| Nemacheilus longistriatus | 109 | 55–82 | 61–78 | 57–68 | 59–75 |
| Nemacheilus pallidus | 2 | 59–76 | |||
| Neodontobutis aurarmus | 3 | 37–37 | 29–30 | ||
| Notopterus | 36 | 280–280 | 281–281 | 193–370 | |
| Ompok bimaculatus | 15 | 65–91 | |||
| Oreochromis niloticus | 21 | 51–313 | 265–265 | 85–173 | 81–285 |
| Osteochilus hasselti | 12 | 61–170 | 78–78 | 134–191 | |
| Osteochilus lini | 27 | 45–95 | 94–120 | 36–155 | 60–84 |
| Osteochilus waandersii | 1 | 134–134 | |||
| Oxyeleotris marmorata | 10 | 30–55 | |||
| Pangasius macronema | 6 | 77–97 | 85–90 | ||
| Parachela siamensis | 326 | 42–135 | 91–142 | 43–125 | 89–136 |
| Parachela spp. | 2024 | 35–95 | 45–68 | 40–75 | 35–76 |
| Parambassis siamensis | 16669 | 25–70 | 30–135 | 28–78 | 38–96 |
| Parasikukia maculata | 13 | 63–63 | 48–65 | ||
| Poropuntius normani | 79 | 35–71 | 25–210 | 37–90 | 48–76 |
| Pristolepis fasciata | 9 | 121–121 | 77–77 | 50–145 | 85–102 |
| Probarbus jullieni | 155 | 83–151 | 83–151 | 80–140 | 80–168 |
| Pseudolais pleurotaenia | 4 | 115–115 | 86–105 | ||
| Puntioplites falcifer | 873 | 40–115 | 54–170 | 41–127 | 43–116 |
| Puntius aurotaeniatus | 6 | 31–55 | |||
| Puntius brevis | 1222 | 22–100 | 41–100 | 25–105 | 41–105 |
| Puntius orphoides | 7 | 75-80 | 78–78 | 70–150 | |
| Puntius partipentazona | 263 | 31–54 | 30–52 | 38–46 | |
| Puntius proctozystron | 170 | 38–106 | 56–132 | 47–115 | 50–110 |
| Puntius stolickzcanus | 1 | 45–45 | |||
| Raiamas guttatus | 39 | 62–135 | 105–128 | 55–168 | 71–126 |
| Rasbora aurotaenia | 1846 | 70–130 | 35–115 | 60–127 | 41–104 |
| Rasbora borapetensis | 286 | 34–59 | 38–60 | 33–61 | 38–58 |
| Rasbora daniconius | 1153 | 37–90 | 58–95 | 51–98 | 51–90 |
| Rasbora dusonensis | 5 | 55–65 | |||
| Rasbora trilineata | 411 | 43–87 | 50–78 | 33–74 | 48–75 |
| Rhinogobius mekongianus | 1 | 68–68 | |||
| Scaphognathops stejnegeri | 43 | 52-95 | 56–86 | 25–81 | 60–80 |
| Sikukia gudgeri | 1640 | 57–100 | 25–146 | 25–102 | 59–126 |
| Tenualosa thibaudeaui | 8 | 113–113 | 85–145 | 74–100 | |
| Thynnichthys thynnoides | 70 | 80–170 | 125–182 | 140–240 | 92–172 |
| Toxotes chatareus | 11 | 82–155 | 95–95 | 86–113 | |
| Trichopodus microlepis | 7 | 56–78 | 111–134 | 102–102 | |
| Trichopodus pectoralis | 2 | 60–60 | 185–185 | ||
| Trichopodus trichopterus | 7 | 49–95 | 75–75 | 91–91 | |
| Trichopsis vittata | 1 | 61–61 | |||
| Unknown 1 | 1 | 99–99 | |||
| Unknown 2 | 1 | 90–90 | |||
| Wallago attu | 2 | 144–147 | |||
| Xenentodon sp. | 1025 | 105–260 | 115–265 | 100–253 | 115–240 |
| Yasuhikotakia lecontei | 30 | 45–66 | 45–66 | 46–62 | 50–52 |
| Yasuhikotakia morleti | 1 | 50–50 | |||
References
- Dugan, P.J.; Barlow, C.; Agostinho, A.A.; Baran, E.; Cada, G.F.; Chen, D.; Cowx, I.G.; Ferguson, J.W.; Jutagate, T.; Mallen-Cooper, M.; et al. Fish migration, dams, and loss of ecosystem services in the Mekong Basin. Ambio 2010, 39, 344–348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winemiller, K.O.; McIntyre, P.B.; Castello, L.; Fluet-Chouinard, E.; Giarrizzo, T.; Nam, S.; Baird, I.G.; Darwall, W.; Lujan, N.K.; Harrison, I.; et al. Balancing hydropower and biodiversity in the Amazon, Congo, and Mekong. Science 2016, 351, 128–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orr, S.; Pittock, J.; Chapagain, A.; Dumaresq, D. Dams on the Mekong River: Lost fish protein and the implications for land and water resources. Glob. Environ. Chang. 2012, 22, 925–932. [Google Scholar] [CrossRef]
- Pringle, C.M.; Freeman, M.C.; Freeman, B.J. Regional effects of hydrologic alterations on riverine macrobiota in the new world: Tropical-temperate comparisons: The massive scope of large dams and other hydrologic modifications in the temperate New World has resulted in distinct regional trends of biotic impoverishment. While neotropical rivers have fewer dams and limited data upon which to make regional generalizations, they are ecologically vulnerable to increasing hydropower development and biotic patterns are emerging. BioScience 2000, 50, 807–823. [Google Scholar]
- Ziv, G.; Baran, E.; Nam, S.; Rodríguez-Iturbe, I.; Levin, S.A. Trading-off fish biodiversity, food security, and hydropower in the Mekong River Basin. Proc. Natl. Acad. Sci. USA 2012, 109, 5609–5614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baumgartner, L.J.; Marsden, T.; Singhanouvong, D.; Phonekhampheng, O.; Stuart, I.G.; Thorncraft, G. Using an experimental in situ fishway to provide key design criteria for lateral fish passage in tropical rivers: A case study from the Mekong River, Central Lao PDR. River Res. Appl. 2012, 28, 1217–1229. [Google Scholar] [CrossRef]
- Stuart, I.G.; Mallen-Cooper, M. An assessment of the effectiveness of a vertical-slot fishway for non-salmonid fish at a tidal barrier on a large tropical/subtropical river. Regul. Rivers Res. Manag. 1999, 15, 575–590. [Google Scholar] [CrossRef]
- Stuart, I.; Berghuis, A. Upstream passage of fish through a vertical-slot fishway in an Australian subtropical river. Fish. Manag. 2002, 9, 111–122. [Google Scholar] [CrossRef]
- Petts, G.E. Impounded Rivers: Perspectives for Ecological Management; Wiley: Chichester, UK, 1984. [Google Scholar]
- Welcomme, R.L. River Fisheries; FAO Fisheries: Rome, Italy, 1985; p. 330. [Google Scholar]
- Baumgartner, L.J.; Boys, C.A.; Marsden, T.; McPherson, J.; Ning, N.; Phonekhampheng, O.; Robinson, W.A.; Singhanouvong, D.; Stuart, I.G.; Thorncraft, G. Comparing fishway designs for application in a large tropical river system. Ecol. Eng. 2018, 120, 36–43. [Google Scholar] [CrossRef]
- Baumgartner, L.J.; Marsden, T.; Millar, J.; Thorncraft, G.; Phonekhampeng, O.; Singhanouvong, D.; Homsombath, K.; Robinson, W.A.; McPherson, J.; Martin, K.; et al. Development of Fish Passage Technology to Increase Fisheries Production on Floodplains in the Lower Mekong Basin; Australian Centre for International Agricultural Research: Canberra, Australia, 2016; 105p. [Google Scholar]
- Marsden, T. Glenore Weir Cone Fishway Monitoring Report; Australasian Fish Passage Services: Fern Bay, Australia, 2017; 29p. [Google Scholar]
- Marsden, T.; Berghuis, A.; Stuart, I. Fitzroy Barrage Cone Fishway Upgrade and Monitoring Report. Report to the Fitzroy Basin Association; Fisheries Collective: Fern Bay, Australia, 2017; 14p. [Google Scholar]
- Stuart, I.G.; Marsden, T.J. Evaluation of cone fishways to facilitate passage of small-bodied fish. Aquac. Fish. 2019. [Google Scholar] [CrossRef]
- Baumgartner, L.J.; Barwick, M.; Boys, C.; Marsden, T.; Martin, K.; McPherson, J.; Ning, N.; Phonekhampeng, O.; Robinson, W.; Singhanouvong, D.; et al. A cautionary tale about the potential impacts of gated culverts on fish passage restoration efforts. J. Ecohydraul. 2019, 4, 27–42. [Google Scholar] [CrossRef]
- Van Zalinge, N.; Degen, P.; Pongsri, C.; Nuov, S.; Jensen, J.; Nguyen, V.; Choulamany, X. The Mekong River System. In Second International Symposium on the Management of Large Rivers for Fisheries; Mekong River Commission: Phnom Penh, Cambodia, 2004; pp. 333–355. [Google Scholar]
- Hortle, K.G. Consumption and Yield of Fish and Other Aquatic Animals from the Lower Mekong Basin; Mekong River Commission: Vientiane, Laos, 2007; p. 87. [Google Scholar]
- Barlow, C.; Baran, E.; Halls, A.S.; Kshatriya, M. How much of the Mekong fish catch is at risk from mainstream dam development? Catch Cult. 2008, 14, 16–21. [Google Scholar]
- Ferguson, J.; Healey, M.; Dugan, P.; Barlow, C. Potential effects of dams on migratory fish in the Mekong River: Lessons from salmon in the Fraser and Columbia rivers. Environ. Manag. 2011, 47, 141–159. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.X.; Siew, R.Y. Water discharge and sediment flux over the past decades in the Lower Mekong River: Possible impacts of the Chinese Dams. Hydrol. Earth Syst. Sci. 2006, 10, 181–195. [Google Scholar] [CrossRef] [Green Version]
- Mekong River Commission. State of the Basin Report; Mekong River Commission: Phnom Penh, Cambodia, 2003; 50p, ISSN 1728:3248. [Google Scholar]
- Kite, G. Modelling the Mekong: Hydrological simulation for environmental impact studies. J. Hydrol. 2001, 253, 1–13. [Google Scholar] [CrossRef]
- Millar, J.; Robinson, W.; Baumgartner, L.; Homsombath, K.; Chittavong, M.; Phommavong, T.; Singhanouvong, D. Local perceptions of changes in the use and management of floodplain fisheries commons: The case of Pak Peung wetland in Lao PDR. Environ. Dev. Sustain. 2019, 21, 1835–1852. [Google Scholar] [CrossRef]
- Mallen-Cooper, M. Developing fishways for non-salmonid fishes: A case study from the Murray River in Australia. In Innovations in Fish Passage, Technology; Odeh, M., Ed.; American Fisheries Society: Bethesda, MD, USA, 1999; pp. 173–195. [Google Scholar]
- Scheffe, H. A method for judging all contrasts in the analysis of variance. Biometrika 1953, 40, 87–110. [Google Scholar]
- Neter, J.; Kutner, M.H.; Nachtsheim, C.J.; Wasserman, W. Applied Linear Statistical Models; WCB McGraw-Hill: New York, NY, USA, 1996; Volume 4. [Google Scholar]
- Mallen-Cooper, M.; Brand, D.M. Non-salmonids in a salmonid fishway: What do 50 years of data tell us about past and future fish passage? Fish. Manag. Ecol. 2007, 14, 319–332. [Google Scholar] [CrossRef]
- Oldani, N.O.; Baigun, C.R.M. Performance of a fishway system in a major South American dam on the Parana River (Argentina-Paraguay). River Res. Appl. 2002, 18, 171–183. [Google Scholar] [CrossRef]
- Anderson, M.J.; Gorley, R.N.; Clarke, K.R. PERMANOVA+ for PRIMER: Guide to Software and Statistical Methods; PRIMER-E: Plymouth, UK, 2008. [Google Scholar]
- Clarke, K.R.; Warwick, R.M. Change in Marine Communities: An Approach to Statistical Analysis and Interpretation, 2nd ed.; Plymouth Marine Laboratory: Plymouth, UK, 2001. [Google Scholar]
- Stuart, I.G.; Zampatti, B.P.; Baumgartner, L.J. Can a low-gradient vertical-slot fishway provide passage for a lowland river fish community? Mar. Freshw. Res. 2008, 59, 332–346. [Google Scholar] [CrossRef]
- Bayley, P.B. Understanding large river-floodplain ecosystems. BioScience 1995, 45, 153–158. [Google Scholar] [CrossRef]
- Marsden, T.; Stuart, I. Fish passage developments for small-bodied tropical fish: Field case-studies lead to technology improvements. J. Ecohydraulics 2019, 4, 14–26. [Google Scholar] [CrossRef]
- Nikora, V.I.; Aberle, J.; Biggs, B.; Jowett, I.; Sykes, J. Effects of fish size, time-to-fatigue and turbulence on swimming performance: A case study of Galaxias maculatus. J. Fish Biol. 2003, 63, 1365–1382. [Google Scholar] [CrossRef]
- Thiem, J.D.; Dawson, J.W.; Hatin, D.; Danylchuk, A.J.; Dumont, P.; Gleiss, A.C.; Wilson, R.P.; Cooke, S.J. Swimming activity and energetic costs of adult lake sturgeon during fishway passage. J. Exp. Biol. 2016, 219, 2534–2544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mallen-Cooper, M.; Zampatti, B.P.; Stuart, I.G.; Baumgartner, L.J. Innovative Fishways—Manipulating Turbulence in the Vertical-Slot Design to Improve Performance and Reduce Cost; Fishway Consulting Services: Sydney, Australia, 2008; p. 19. [Google Scholar]
- Foulds, W.L.; Lucas, M.C. Extreme inefficiency of two conventional, technical fishways used by European river lamprey (Lampetra fluviatilis). Ecol. Eng. 2013, 58, 423–433. [Google Scholar] [CrossRef] [Green Version]
- Baumgartner, L.J.; Stuart, I.G.; Zampatti, B.P. Determining diel variation in fish assemblages downstream of three weirs in a regulated lowland river. J. Fish Biol. 2008, 72, 218–232. [Google Scholar] [CrossRef]
- Nunn, A.D.; Copp, G.H.; Vilizzi, L.; Carter, M.G. Seasonal and diel patterns in the migrations of fishes between a river and a floodplain tributary. Ecol. Freshw. Fish 2010, 19, 153–162. [Google Scholar] [CrossRef]
- Castro-Santos, T.; Haro, A.; Walk, S. A passive integrated transponder (PIT) tag system for monitoring fishways. Fish. Res. 1996, 28, 253–261. [Google Scholar] [CrossRef]
- Dudgeon, D. Asian river fishes in the Anthropocene: Threats and conservation challenges in an era of rapid environmental change. J. Fish Biol. 2011, 79, 1487–1524. [Google Scholar] [CrossRef]






| Species | Entrance | Exit | Dissimilarity Contribution (%) |
|---|---|---|---|
| Rasbora daniconius | 0.72 | 0.17 | 3.04 |
| Hypsibarbus lagleri | 0.33 | 0.78 | 2.9 |
| Amblyrhynchichthys micracanthus | 0.33 | 0.72 | 2.84 |
| Barbonymus altus | 0.39 | 0.67 | 2.66 |
| Puntius partipentazona | 0.61 | 0 | 2.62 |
| Parachela spp. | 0.56 | 0.17 | 2.4 |
| Species | Entrance | Exit | Dissimilarity Contribution (%) |
|---|---|---|---|
| Poropuntius normani | 0.65 | 0.18 | 2.44 |
| Rasbora borapetensis | 0.65 | 0.29 | 2.42 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baumgartner, L.J.; Boys, C.; Marsden, T.; McPherson, J.; Ning, N.; Phonekhampheng, O.; Robinson, W.; Singhanouvong, D.; Stuart, I.G.; Thorncraft, G. A Cone Fishway Facilitates Lateral Migrations of Tropical River-Floodplain Fish Communities. Water 2020, 12, 513. https://doi.org/10.3390/w12020513
Baumgartner LJ, Boys C, Marsden T, McPherson J, Ning N, Phonekhampheng O, Robinson W, Singhanouvong D, Stuart IG, Thorncraft G. A Cone Fishway Facilitates Lateral Migrations of Tropical River-Floodplain Fish Communities. Water. 2020; 12(2):513. https://doi.org/10.3390/w12020513
Chicago/Turabian StyleBaumgartner, Lee J., Craig Boys, Tim Marsden, Jarrod McPherson, Nathan Ning, Oudom Phonekhampheng, Wayne Robinson, Douangkham Singhanouvong, Ivor G. Stuart, and Garry Thorncraft. 2020. "A Cone Fishway Facilitates Lateral Migrations of Tropical River-Floodplain Fish Communities" Water 12, no. 2: 513. https://doi.org/10.3390/w12020513