Migration of Non-Native Predator Asp (Leuciscus aspius) from a Reservoir Poses a Potential Threat to Native Species in Tributaries


Abstract
:1. Introduction
2. Materials and Methods
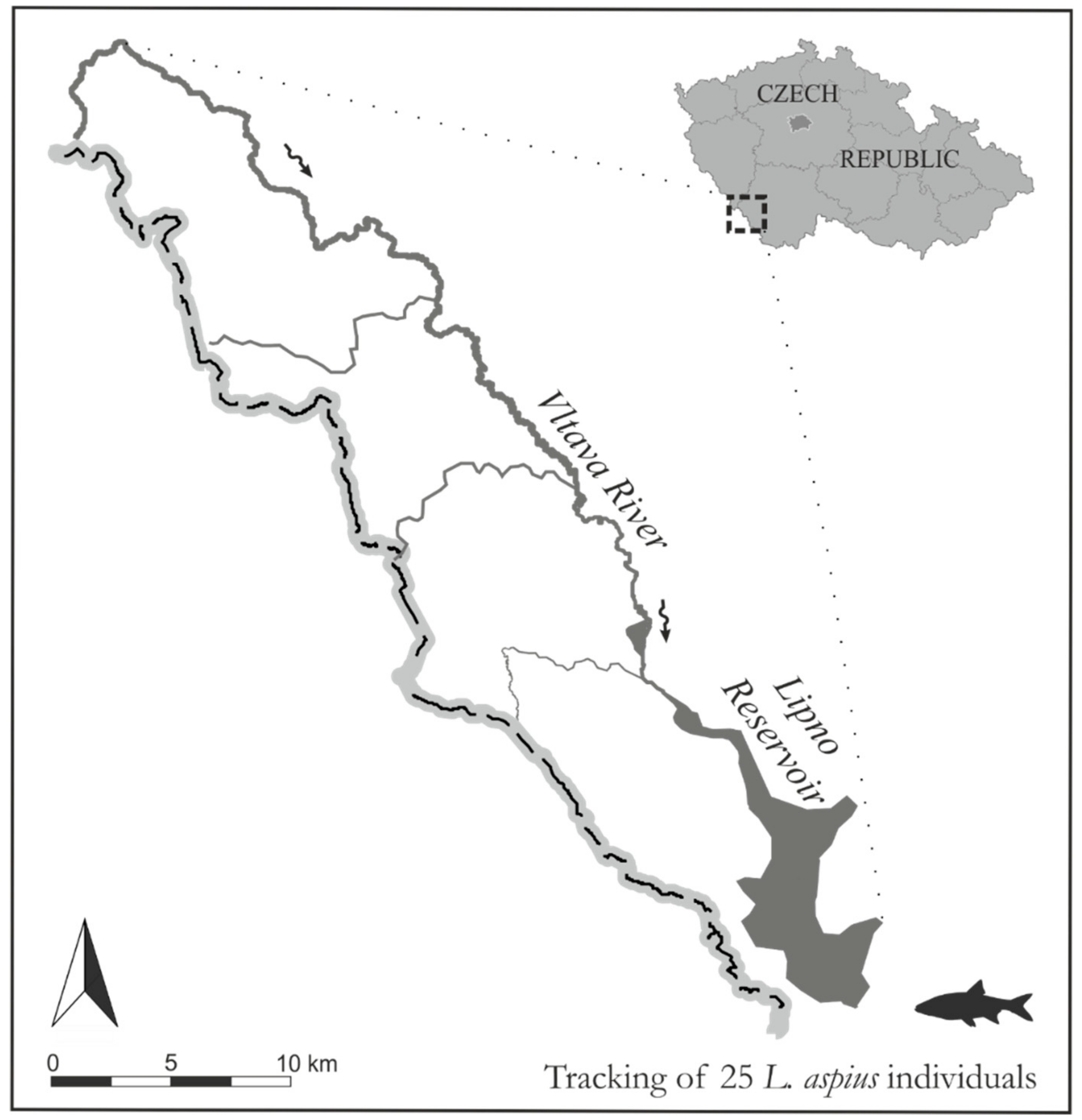
2.1. Study Area
2.2. Fish Origin and Tagging
2.3. Monitoring Procedures
2.4. Habitat Measurements
2.5. Data Analyses
2.6. Statistical Analysis
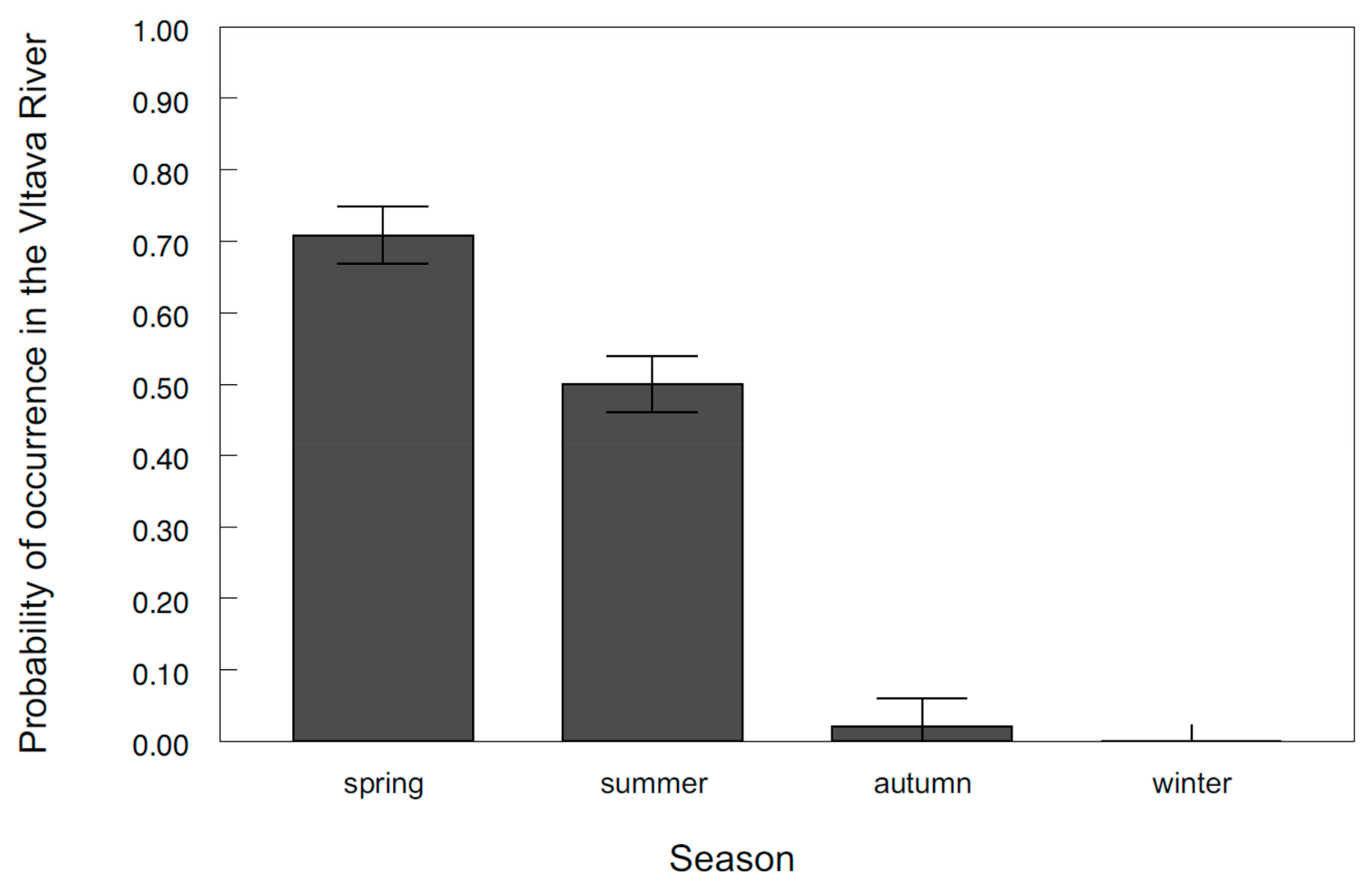
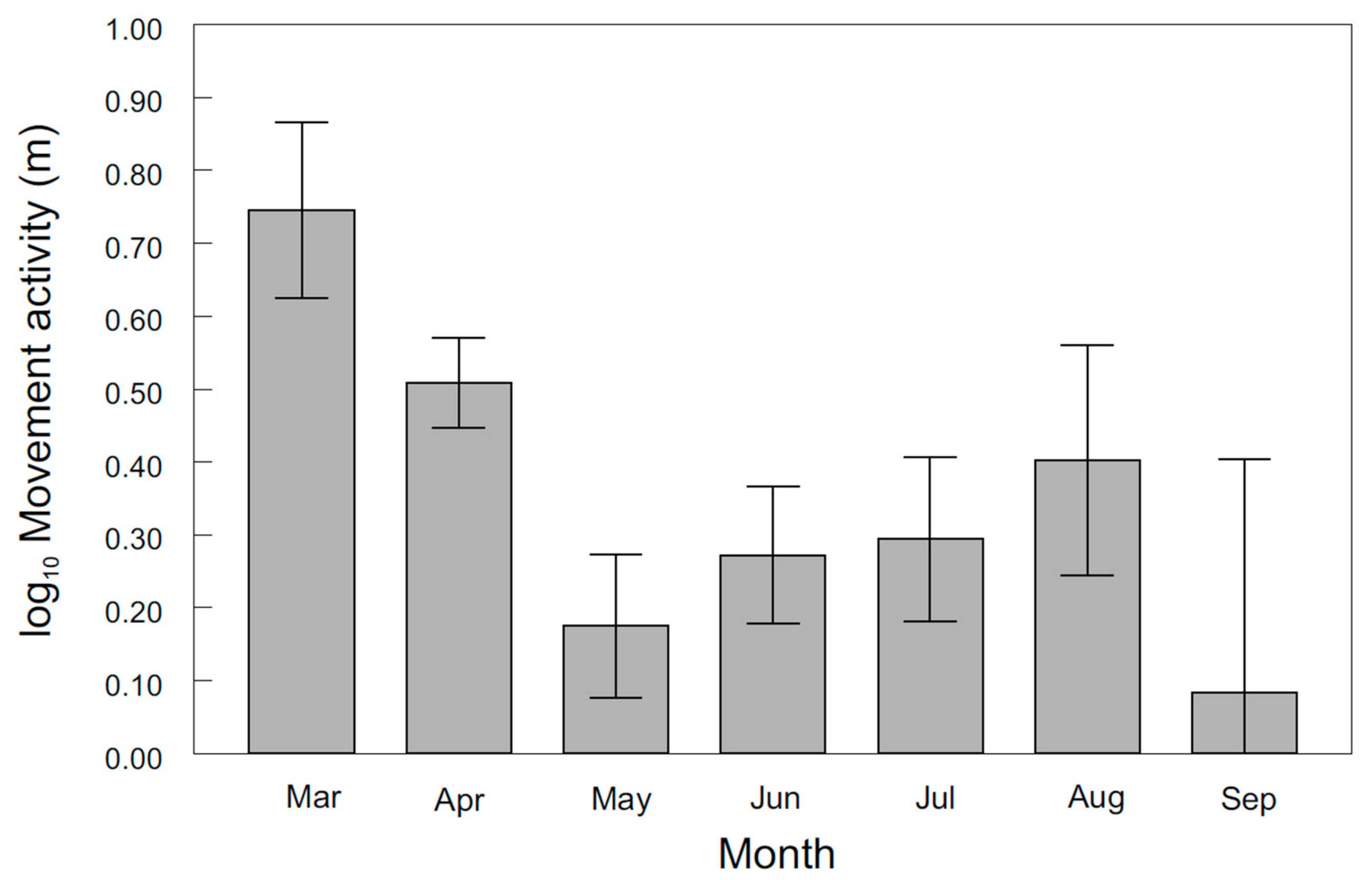
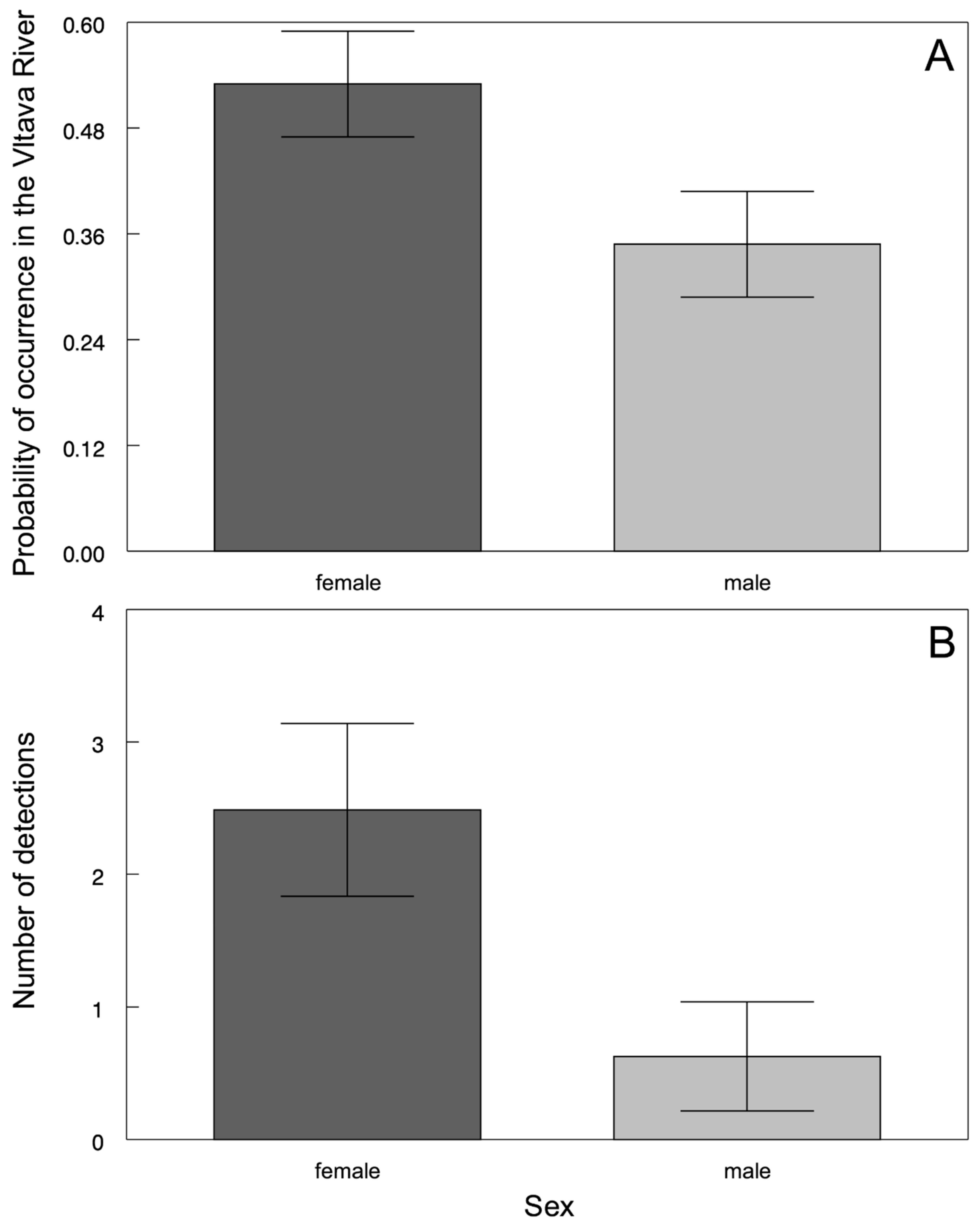
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sala, O.E.; Chapin, F.S.; Armesto, J.J.; Berlow, E.; Bloomfield, J.; Dirzo, R.; Huber-Sanwald, E.; Huenneke, L.F.; Jackson, R.B.; Kinzig, A.; et al. Global biodiversity scenarios for the year 2100. Science 2000, 287, 1770–1774. [Google Scholar] [CrossRef]
- Dudgeon, D.; Arthington, A.H.; Gessner, M.O.; Kawabata, Z.I.; Knowler, D.J.; Lévêque, C.; Naiman, R.J.; Prieur-Richard, A.H.; Soto, D.; Stiassny, M.L.J.; et al. Freshwater biodiversity: Importance, threats, status and conservation challenges. Biol. Rev. Camb. Philos. Soc. 2006, 81, 163–182. [Google Scholar] [CrossRef] [PubMed]
- Kiruba-Sankar, R.; Velmurugan, A.; Saravanan, K.; Dam Roy, S.; Praveen Raj, J.; Raymond Jani Angel, J.; Lohith Kumar, K. Invasive Species in Freshwater Ecosystems—Threats to Ecosystem Services. In Biodiversity and Climate Change Adaptation in Tropical Islands; Sivaperuman, C., Velmurugan, A., Singh, A., Jaisankar, I., Eds.; Elsevier Inc.: Amsterdam, The Netherlands, 2018; pp. 257–296. ISBN 9780128130643. [Google Scholar]
- Mack, R.N.; Simberloff, D.; Lonsdale, W.M.; Evans, H.; Clout, M.; Bazzaz, F.A. Biotic invasions: Causes, epidemiology, global consequences, and control. Ecol. Appl. 2000, 10, 689–710. [Google Scholar] [CrossRef]
- Gozlan, R.E.; Britton, J.R.; Cowx, I.; Copp, G.H. Current knowledge on non-native freshwater fish introductions. J. Fish Biol. 2010, 76, 751–786. [Google Scholar] [CrossRef]
- Roy, H.E.; Bacher, S.; Essl, F.; Adriaens, T.; Aldridge, D.C.; Bishop, J.D.D.; Blackburn, T.M.; Branquart, E.; Brodie, J.; Carboneras, C.; et al. Developing a list of invasive alien species likely to threaten biodiversity and ecosystems in the European Union. Glob. Chang. Biol. 2019, 25, 1032–1048. [Google Scholar] [CrossRef] [PubMed]
- Lodge, D.M. Biological invasions: Lessons for ecology. Trends Ecol. Evol. 1993, 8, 133–137. [Google Scholar] [CrossRef]
- Vitousek, P.M.; Mooney, H.A.; Lubchenco, J.; Melillo, J.M. Human Domination of Earth’s Ecosystems. In Urban Ecology: An International Perspective on the Interaction Between Humans and Nature; Springer: New York, NY, USA, 2008; pp. 3–13. ISBN 9780387734118. [Google Scholar]
- Asche, F.; Roll, K.H.; Sandvold, H.N.; Sørvig, A.; Zhang, D. Salmon aquaculture: Larger companies and increased production. Aquac. Econ. Manag. 2013, 17, 322–339. [Google Scholar] [CrossRef]
- Copp, G.H.; Bianco, P.G.; Bogutskaya, N.G.; Eros, T.; Falka, I.; Ferreira, M.T.; Fox, M.G.; Freyhof, J.; Gozlan, R.E.; Grabowska, J.; et al. To be, or not to be, a non-native freshwater fish? J. Appl. Ichthyol. 2005, 21, 242–262. [Google Scholar] [CrossRef]
- Casal, C.M.V. Global documentation of fish introductions: The growing crisis and recommendations for action. Biol. Invasions 2006, 8, 3–11. [Google Scholar] [CrossRef]
- Carosi, A.; Ghetti, L.; La Porta, G.; Lorenzoni, M. Ecological effects of the European barbel Barbus barbus (L., 1758) (Cyprinidae) invasion on native barbel populations in the Tiber River basin (Italy). Eur. Zool. J. 2017, 84, 420–435. [Google Scholar] [CrossRef]
- Cook, E.J.; Ashton, G.; Campbell, M.; Coutts, A.; Gollasch, S.; Hewitt, C.; Liu, H.; Minchin, D.; Ruiz, G.; Shucksmith, R. Non-Native Aquaculture Species Releases: Implications for Aquatic Ecosystems. In Aquaculture in the Ecosystem; Springer: Dordrecht, The Netherlands, 2008; pp. 155–184. ISBN 9781402068102. [Google Scholar]
- Welcomme, R.L. A history of international introduction of inland aquatic species. ICES Mar. Sci. Symp. 1992, 194, 3–14. [Google Scholar]
- Azevedo-Santos, V.M.; Vitule, J.R.S.; Pelicice, F.M.; García-Berthou, E.; Simberloff, D. Nonnative fish to control aedes mosquitoes: A controversial, harmful tool. Bioscience 2017, 67, 84–90. [Google Scholar] [CrossRef]
- Pyke, G.H. Plague Minnow or Mosquito fish? A review of the biology and impacts of introduced Gambusia species. Annu. Rev. Ecol. Evol. Syst. 2008, 39, 171–191. [Google Scholar] [CrossRef]
- Cucherousset, J.; Horky, P.; Slavík, O.; Ovidio, M.; Arlinghaus, R.; Boulêtreau, S.; Britton, R.; García-Berthou, E.; Santoul, F. Ecology, behaviour and management of the European catfish. Rev. Fish Biol. Fish. 2018, 28, 177–190. [Google Scholar] [CrossRef]
- Copp, G.G.H.; Robert Britton, J.; Cucherousset, J.; García-Berthou, E.; Kirk, R.; Peeler, E.; Stakėnas, S.; Britton, J.R.; Cucherousset, J. Voracious invader or benign feline? A review of the environmental biology of European catfish Silurus glanis in its native and introduced ranges. Fish Fish. 2009, 10, 252–282. [Google Scholar] [CrossRef]
- Gherardi, F. Biological Invasions in Inland Waters: An Overview. In Biological Invaders in Inland Waters: Profiles, Distribution, and Threats; Gherardi, F., Ed.; Springer: Dordrecht, The Netherlands, 2007; pp. 3–25. ISBN 978-1-4020-6029-8. [Google Scholar]
- Boulêtreau, S.; Gaillagot, A.; Carry, L.; Tétard, S.; De Oliveira, E.; Santoul, F. Adult Atlantic salmon have a new freshwater predator. PLoS ONE 2018, 13, e0196046. [Google Scholar] [CrossRef]
- Perry, A.L.; Low, P.J.; Ellis, J.R.; Reynolds, J.D. Ecology: Climate change and distribution shifts in marine fishes. Science 2005, 308, 1912–1915. [Google Scholar] [CrossRef] [PubMed]
- Britton, J.R.; Cucherousset, J.; Davies, G.D.; Godard, M.J.; Copp, G.H. Non-native fishes and climate change: Predicting species responses to warming temperatures in a temperate region. Freshw. Biol. 2010, 55, 1130–1141. [Google Scholar] [CrossRef]
- Pederson, J.; Mieszkowska, N.; Carlton, J.T.; Gollasch, S.; Jelmert, A.; Minchin, D.; Occhipinti-Ambrogi, A.; Wallentinus, I. Climate Change and Non-Native Species in the North Atlantic. In ICES Status Report on Climate Change in the North Atlantic; ICES Cooperative Research Report No. 310; ICES: Copenhagen, Denmark, 2011; pp. 174–190. [Google Scholar]
- Kaukorantana, M.T.; Pennanen, J. Propagation and management of the asp, Aspius aspius (L.), in Finland. In Proceedings of the Management of Freshwater Fisheries: A symposium organized by the European Inland Fisheries Advisory Commission, Goteborg, Sweden, 31 May–3 June 1988; Vandensen, W., Steinmetz, B., Hughes, R., Eds.; Pudoc: Wageningen, The Netherlands, 1990; pp. 67–73. [Google Scholar]
- Radinger, J.; Essl, F.; Hölker, F.; Horký, P.; Slavík, O.; Wolter, C. The future distribution of river fish: The complex interplay of climate and land use changes, species dispersal and movement barriers. Glob. Chang. Biol. 2017, 23, 4970–4986. [Google Scholar] [CrossRef]
- Benitez, J.P.; Dierckx, A.; Nzau Matondo, B.; Rollin, X.; Ovidio, M. Movement behaviours of potamodromous fish within a large anthropised river after the reestablishment of the longitudinal connectivity. Fish. Res. 2018, 207, 140–149. [Google Scholar] [CrossRef]
- Musil, J.; Horký, P.; Slavík, O.; Zbořil, A.; Horká, P. The response of the young of the year fish to river obstacles: Functional and numerical linkages between dams, weirs, fish habitat guilds and biotic integrity across large spatial scale. Ecol. Indic. 2012, 23, 634–640. [Google Scholar] [CrossRef]
- Liew, J.H.; Tan, H.H.; Yeo, D.C.J. Dammed rivers: Impoundments facilitate fish invasions. Freshw. Biol. 2016, 61, 1421–1429. [Google Scholar] [CrossRef]
- Vostradovský, J. On the biology of asp (Aspius aspius L.) in the water-supply reservoir Švihov (Želivka). Anim. Husb. 1974, 19, 683–688. [Google Scholar]
- L’Abée-Lund, J.H.; Vøllestad, L.A. Feeding migration of roach, Rutilus rutilus (L.), in Lake Arungen, Norway. J. Fish Biol. 1987, 30, 349–355. [Google Scholar] [CrossRef]
- Hladík, M.; Kubečka, J. Fish migration between a temperate reservoir and its main tributary. Hydrobiologia 2003, 504, 251–266. [Google Scholar] [CrossRef]
- Hladík, M.; Kubečka, J.; Mrkvička, T.; Čech, M.; Draštík, V.; Frouzová, J.; Hohausová, E.; Matěna, J.; Matěnová, V.; Kratochvíl, M.; et al. Effects of the construction of a reservoir on the fish assemblage in an inflow river. Czech J. Anim. Sci. 2008, 53, 537–547. [Google Scholar] [CrossRef] [Green Version]
- Lucas, M.C.; Baras, E. Methods for studying spatial behaviour of freshwater fishes in the natural environment. Fish Fish. 2003, 1, 283–316. [Google Scholar] [CrossRef]
- Benitez, J.P.; Nzau Matondo, B.; Dierckx, A.; Ovidio, M. An overview of potamodromous fish upstream movements in medium-sized rivers, by means of fish passes monitoring. Aquat. Ecol. 2015, 49, 481–497. [Google Scholar] [CrossRef]
- Benitez, J.P.; Ovidio, M. The influence of environmental factors on the upstream movements of rheophilic cyprinids according to their position in a river basin. Ecol. Freshw. Fish 2018, 27, 660–671. [Google Scholar] [CrossRef]
- Fredrich, F. Long-term investigations of migratory behaviour of asp (Aspius aspius L.) in the middle part of the Elbe River, Germany. J. Appl. Ichthyol. 2003, 19, 294–302. [Google Scholar] [CrossRef]
- Kulíšková, P.; Horký, P.; Slavík, O.; Jones, J.I. Factors influencing movement behaviour and home range size in ide Leuciscus idus. J. Fish Biol. 2009, 74, 1269–1279. [Google Scholar] [CrossRef] [PubMed]
- Šmejkal, M.; Baran, R.; Blabolil, P.; Vejřík, L.; Prchalová, M.; Vebrová, L.; Ricard, D.; Kubečka, J.; Sajdlová, Z.; Mrkvička, T.; et al. Seasonal and daily protandry in a cyprinid fish. Sci. Rep. 2017, 7, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kottelat, M.; Freyhof, J.J. Handbook of European Freshwater Fishes; IUCN Publication: Gland, Switzerland, 2007; ISBN 978-2-8399-0298-4. [Google Scholar]
- Horký, P.; Slavík, O. Diel and seasonal rhythms of asp Leuciscus aspius (L.) in a riverine environment. Ethol. Ecol. Evol. 2017, 29, 449–459. [Google Scholar] [CrossRef]
- Winter, J.D. Underwater Biotelemetry. In Fisheries Techniques; Nielson, Larry A., Johnson, D.L., Eds.; American Fisheries Society: Bethesda, MD, USA, 1983; pp. 371–395. [Google Scholar]
- SAS Institute Inc. SAS/STAT® 9.1 User’s Guide; SAS Institute Inc.: Cary, NC, USA, 2004; ISBN 1-59047-243-8. [Google Scholar]
- Breslow, N.E.; Clayton, D.G. Approximate Inference in Generalized Linear Mixed Models. J. Am. Stat. Assoc. 1993, 88, 9. [Google Scholar]
- Littell, R.C.; Milliken, G.A.; Stroup, W.W.; Wolfinger, R.D.; Schabenberger, O. SAS® for Mixed Models, Second Edition; SAS Institute Inc.: Cary, NC, USA, 2006; ISBN 1-59047-500-3. [Google Scholar]
- Kenward, M.G.; Roger, J.H. Small sample inference for fixed effects from restricted maximum likelihood. Biometrics 1997, 53, 983–997. [Google Scholar] [CrossRef] [PubMed]
- Liang, K.Y.; Zeger, S.L. Longitudinal data analysis using generalized linear models. Biometrika 1986, 73, 13–22. [Google Scholar] [CrossRef]
- Cucherousset, J.; Olden, J.D. Ecological impacts of non-native freshwater fishes. Fisheries 2011, 36, 215–230. [Google Scholar] [CrossRef]
- Goudswaard, K.; Witte, F.; Katunzi, E.F.B. The invasion of an introduced predator, Nile perch (Lates niloticus, L.) in Lake Victoria (East Africa): Chronology and causes. Environ. Biol. Fishes 2008, 81, 127–139. [Google Scholar] [CrossRef]
- Vašek, M.; Matěna, J.; Říha, M.; Jůza, T.; Peterka, J.; Vejříková, I.; Blabolil, P.; Eloranta, A.P.; Kubečka, J.; Šmejkal, M. Stable isotopes and gut contents indicate differential resource use by coexisting asp (Leuciscus aspius) and pikeperch (Sander lucioperca). Ecol. Freshw. Fish 2018, 27, 1054–1065. [Google Scholar] [CrossRef]
- Brodersen, J.; Nicolle, A.; Nilsson, P.A.; Skov, C.; Brönmark, C.; Hansson, L.A. Interplay between temperature, fish partial migration and trophic dynamics. Oikos 2011, 120, 1838–1846. [Google Scholar] [CrossRef]
- Chapman, B.B.; Skov, C.; Hulthén, K.; Brodersen, J.; Nilsson, P.A.; Hansson, L.-A.; Brönmark, C. Partial migration in fishes: definitions, methodologies and taxonomic distribution. J. Fish Biol. 2012, 81, 479–499. [Google Scholar] [CrossRef] [PubMed]
- Brönmark, C.; Skov, C.; Brodersen, J.; Nilsson, P.A.; Hansson, L.A. Seasonal migration determined by a trade-off between predator avoidance and growth. PLoS ONE 2008, 3, e1957. [Google Scholar] [CrossRef] [PubMed]
- Skov, C.; Baktoft, H.; Brodersen, J.; Brönmark, C.; Chapman, B.B.; Hansson, L.A.; Anders Nilsson, P. Sizing up your enemy: Individual predation vulnerability predicts migratory probability. Proc. R. Soc. B Biol. Sci. 2011, 278, 1414–1418. [Google Scholar] [CrossRef] [PubMed]
- Chapman, B.B.; Hulthén, K.; Blomqvist, D.R.; Hansson, L.A.; Nilsson, J.Å.; Brodersen, J.; Anders Nilsson, P.; Skov, C.; Brönmark, C. To boldly go: Individual differences in boldness influence migratory tendency. Ecol. Lett. 2011, 14, 871–876. [Google Scholar] [CrossRef] [PubMed]
- Brönmark, C.; Nilsson, P.A.; Skov, C.; Hulthén, K.; Chapman, B.B.; Brodersen, J.; Hansson, L.-A. There and back again: migration in freshwater fishes. Can. J. Zool. 2013, 92, 467–479. [Google Scholar] [CrossRef]
- Krpo-Ćetković, J.; Hegediš, A.; Lenhardt, M. Diet and growth of asp, Aspius aspius (Linnaeus, 1758), in the Danube River near the confluence with the Sava River (Serbia). J. Appl. Ichthyol. 2010, 26, 513–521. [Google Scholar] [CrossRef]
- Flecker, A.S.; Townsend, C.R. Community-wide consequences of trout introduction in New Zealand streams. Ecol. Appl. 1994, 4, 798–807. [Google Scholar] [CrossRef]
- Baxter, C.V.; Fausch, K.D.; Murakami, M.; Chapman, P.L. Fish invasion restructures stream and forest Food webs by interrupting reciprocal prey subsidies. Ecology 2004, 85, 2656–2663. [Google Scholar] [CrossRef]
- Mobley, K.B.; Granroth-Wilding, H.; Ellmen, M.; Vähä, J.-P.; Aykanat, T.; Johnston, S.E.; Orell, P.; Erkinaro, J.; Primmer, C.R. Home ground advantage: Local Atlantic salmon have higher reproductive fitness than dispersers in the wild. Sci. Adv. 2019, 5, eaav1112. [Google Scholar] [CrossRef] [Green Version]
- Lucas, M.C.; Baras, E. Migration of Freshwater Fishes; Blackwell Science: Hoboken, NJ, USA, 2001; ISBN 0632057548. [Google Scholar]
- Smith, R.J.F. Social Behaviour, Homing and Migration. In Cyprinid Fishes: Systematics, Biology and Exploitation; Winfield, I.J., Nelson, J.S., Eds.; Springer: Dordrecht, The Netherlands, 1991; pp. 509–529. ISBN 978-94-011-3092-9. [Google Scholar]
- L’Abáe-Lund, J.H.; Vøllestad, L.A. Homing precision of roach Rutilus rutilus in Lake Arungen, Norway. Environ. Biol. Fishes 1985, 13, 235–239. [Google Scholar] [CrossRef]
- Liu, J.K.; Yu, Z.T. Water quality changes and effects on fish populations in the Hanjiang River, China, following hydroelectric dam construction. Regul. Rivers Res. Manag. 1992, 7, 359–368. [Google Scholar] [CrossRef]
- Irving, D.B.; Modde, T. Home-range fidelity and use of historic habitat by adult Colorado pikeminnow (Ptychocheilus lucius) in the White River, Colorado and Utah. West. North Am. Nat. 2000, 60, 16–25. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fish ID | Sex | Standard Length (mm) | Body Size (g) | Number of Detections | Number of Returns |
|---|---|---|---|---|---|
| 3 | male | 480 | 1635 | 4 | 2 |
| 6 | female | 680 | 5620 | 8 | 0 |
| 7 | male | 560 | 2570 | 0 | 0 |
| 8 | male | 650 | 3990 | 4 | 1 |
| 11 | male | 510 | 1975 | 4 | 2 |
| 12 | male | 610 | 3340 | 8 | 0 |
| 13 | male | 660 | 4630 | 0 | 0 |
| 16 | male | 540 | 2240 | 0 | 0 |
| 23 | female | 460 | 1745 | 0 | 0 |
| 24 | male | 575 | 2675 | 6 | 2 |
| 32 | female | 455 | 1255 | 13 | 2 |
| 36 | male | 480 | 1930 | 0 | 0 |
| 37 | male | 450 | 1100 | 24 | 2 |
| 39 | male | 450 | 1350 | 18 | 4 |
| 57 | female | 580 | 2800 | 46 | 4 |
| 59 | female | 470 | 1565 | 26 | 3 |
| 60 | male | 385 | 1205 | 18 | 4 |
| 63 | female | 465 | 1360 | 24 | 2 |
| 140 | male | 530 | 2255 | 1 | 0 |
| 141 | male | 465 | 1540 | 0 | 0 |
| 142 | male | 530 | 2160 | 0 | 0 |
| 146 | male | 470 | 1535 | 0 | 0 |
| 150 | male | 465 | 1470 | 1 | 0 |
| 151 | male | 480 | 1540 | 1 | 0 |
| 152 | male | 520 | 1935 | 0 | 0 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pfauserová, N.; Slavík, O.; Horký, P.; Kolářová, J.; Randák, T. Migration of Non-Native Predator Asp (Leuciscus aspius) from a Reservoir Poses a Potential Threat to Native Species in Tributaries. Water 2019, 11, 1306. https://doi.org/10.3390/w11061306
Pfauserová N, Slavík O, Horký P, Kolářová J, Randák T. Migration of Non-Native Predator Asp (Leuciscus aspius) from a Reservoir Poses a Potential Threat to Native Species in Tributaries. Water. 2019; 11(6):1306. https://doi.org/10.3390/w11061306
Chicago/Turabian StylePfauserová, Nikola, Ondřej Slavík, Pavel Horký, Jitka Kolářová, and Tomáš Randák. 2019. "Migration of Non-Native Predator Asp (Leuciscus aspius) from a Reservoir Poses a Potential Threat to Native Species in Tributaries" Water 11, no. 6: 1306. https://doi.org/10.3390/w11061306