New Methods for Microbiological Monitoring at Riverbank Filtration Sites


Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description
2.2. Sample Collection and Microbiological Characterization
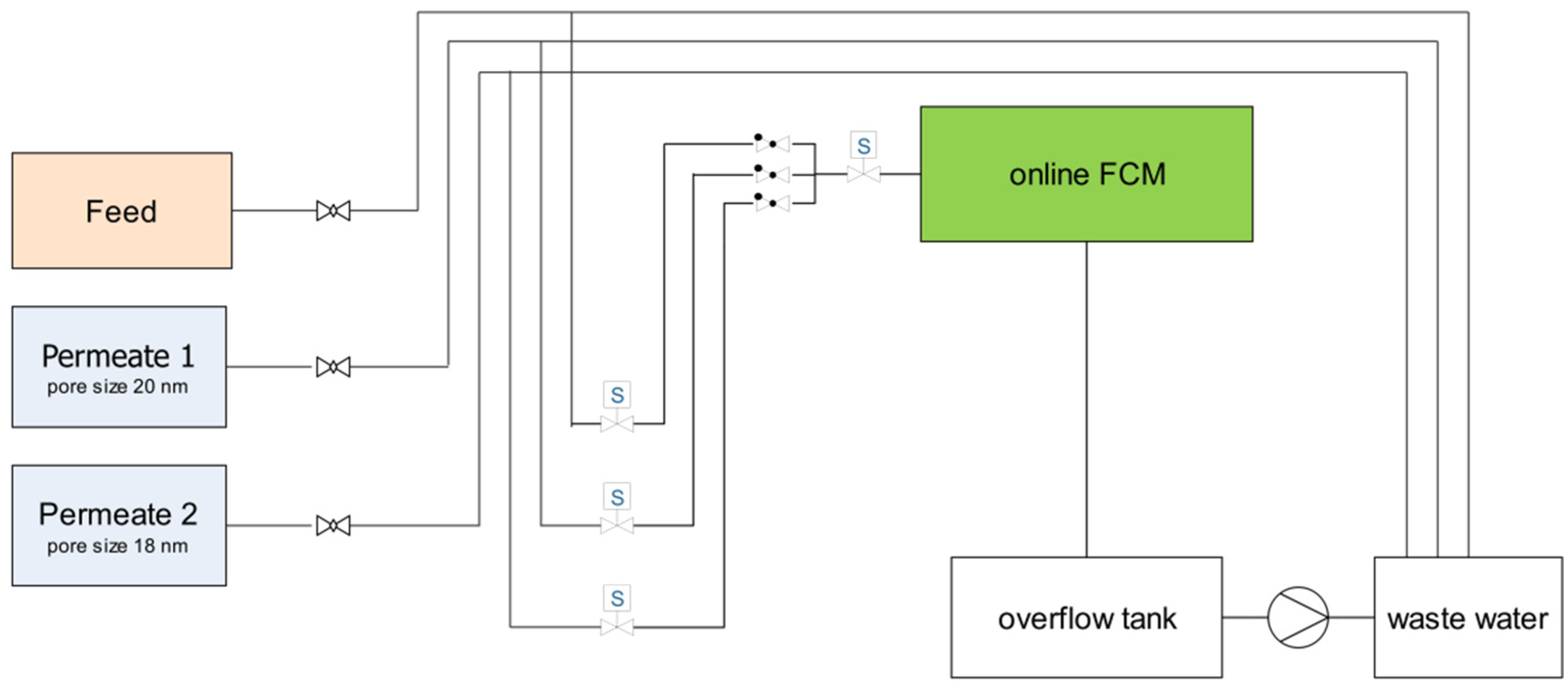
2.3. Ultrafiltration Pilot Plant and Online Flow-Cytometry (FCM) Measurements
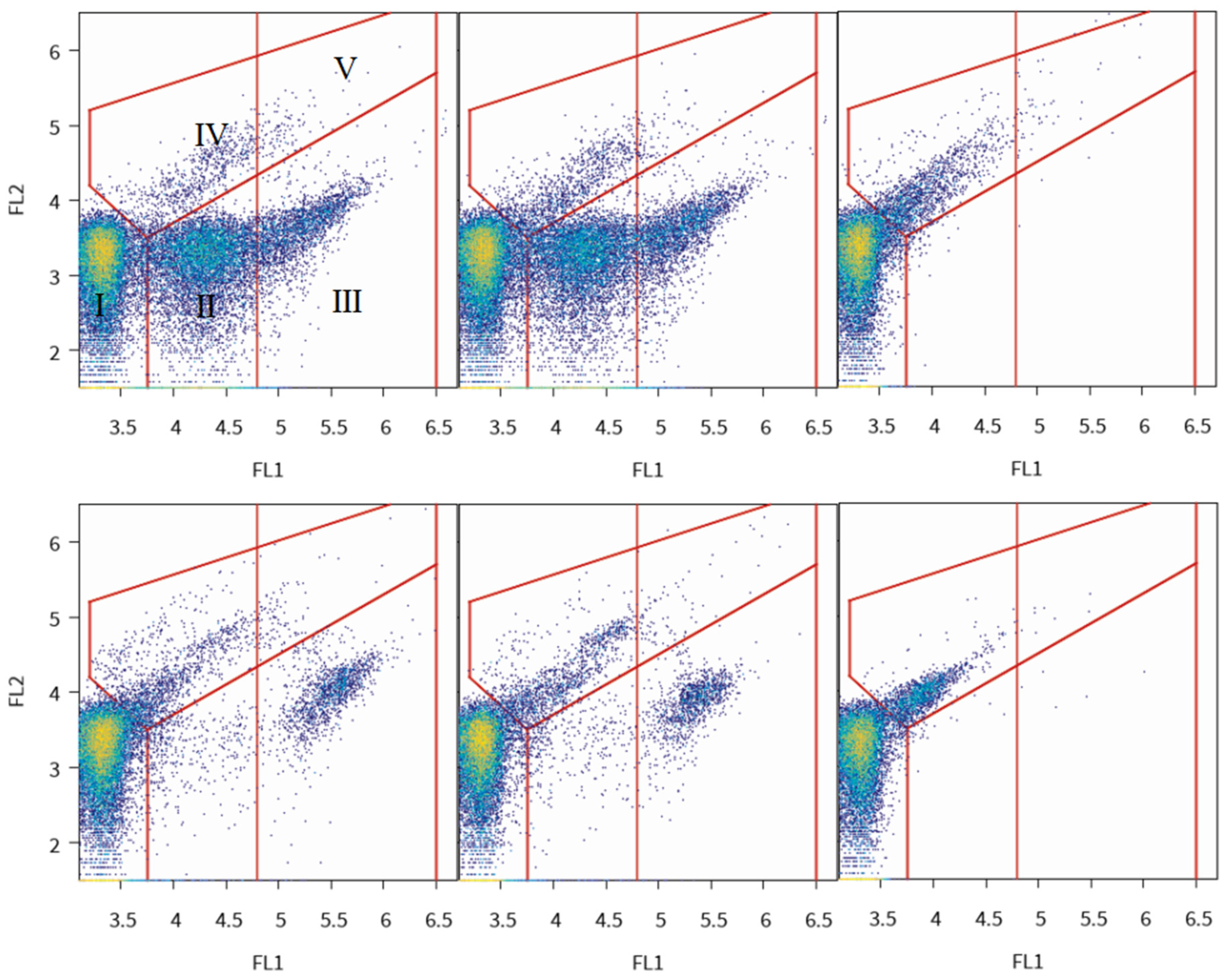
2.4. Data Analysis
3. Results
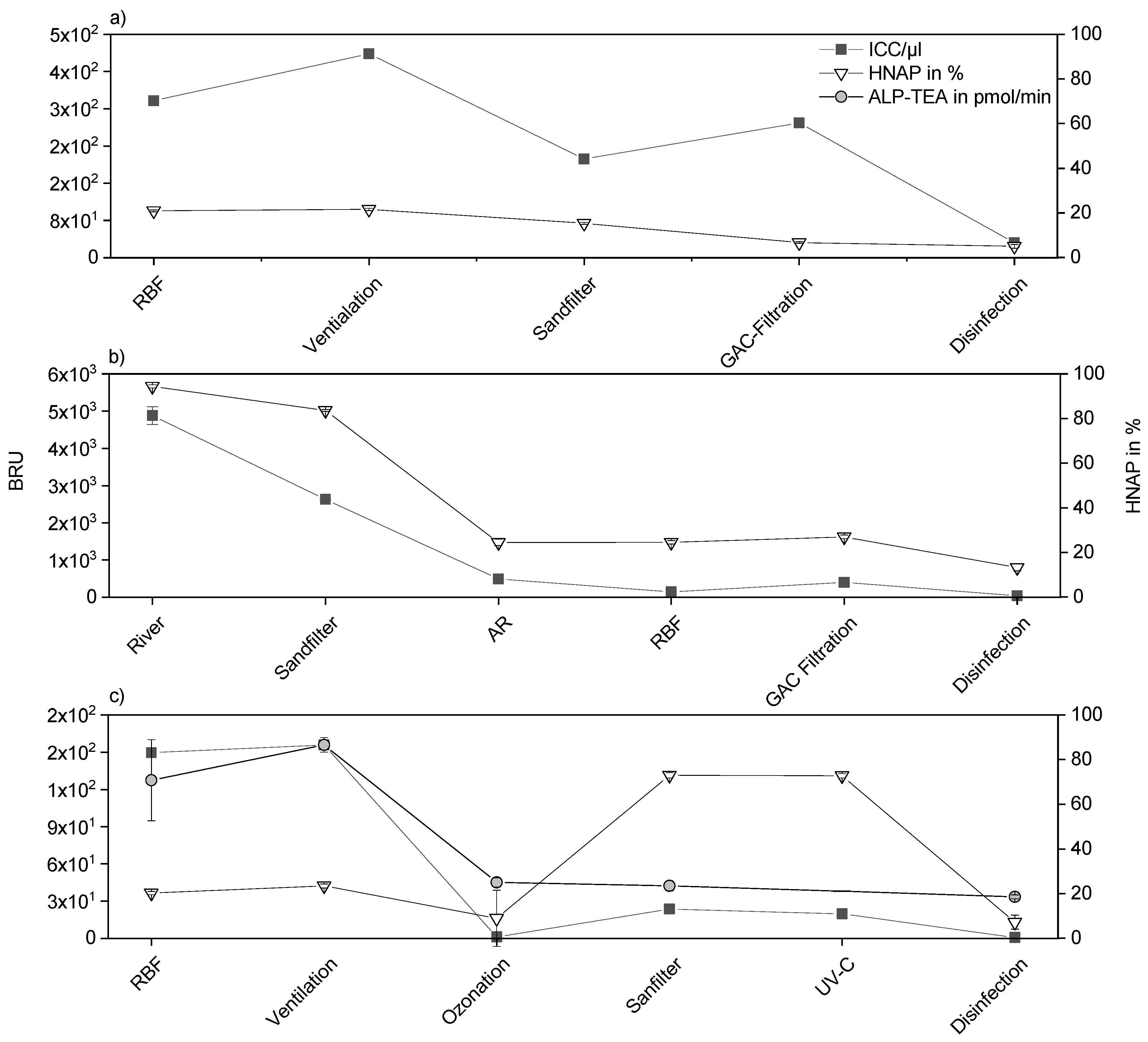
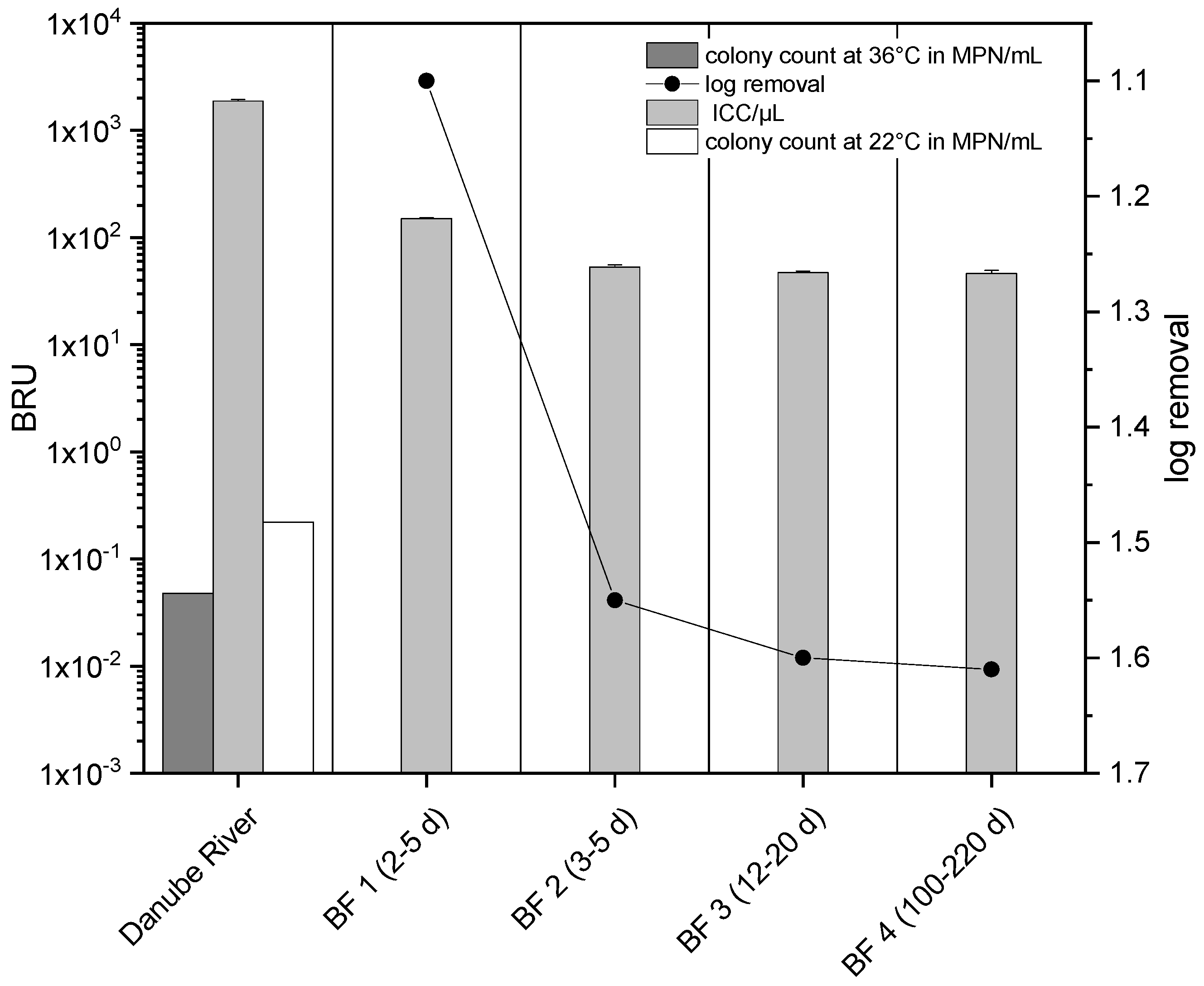
3.1. Correlation of New Methods and Conventional Cultivation-Based Methods during Riverbank Filtration (RBF) and Drinking-Water Treatment
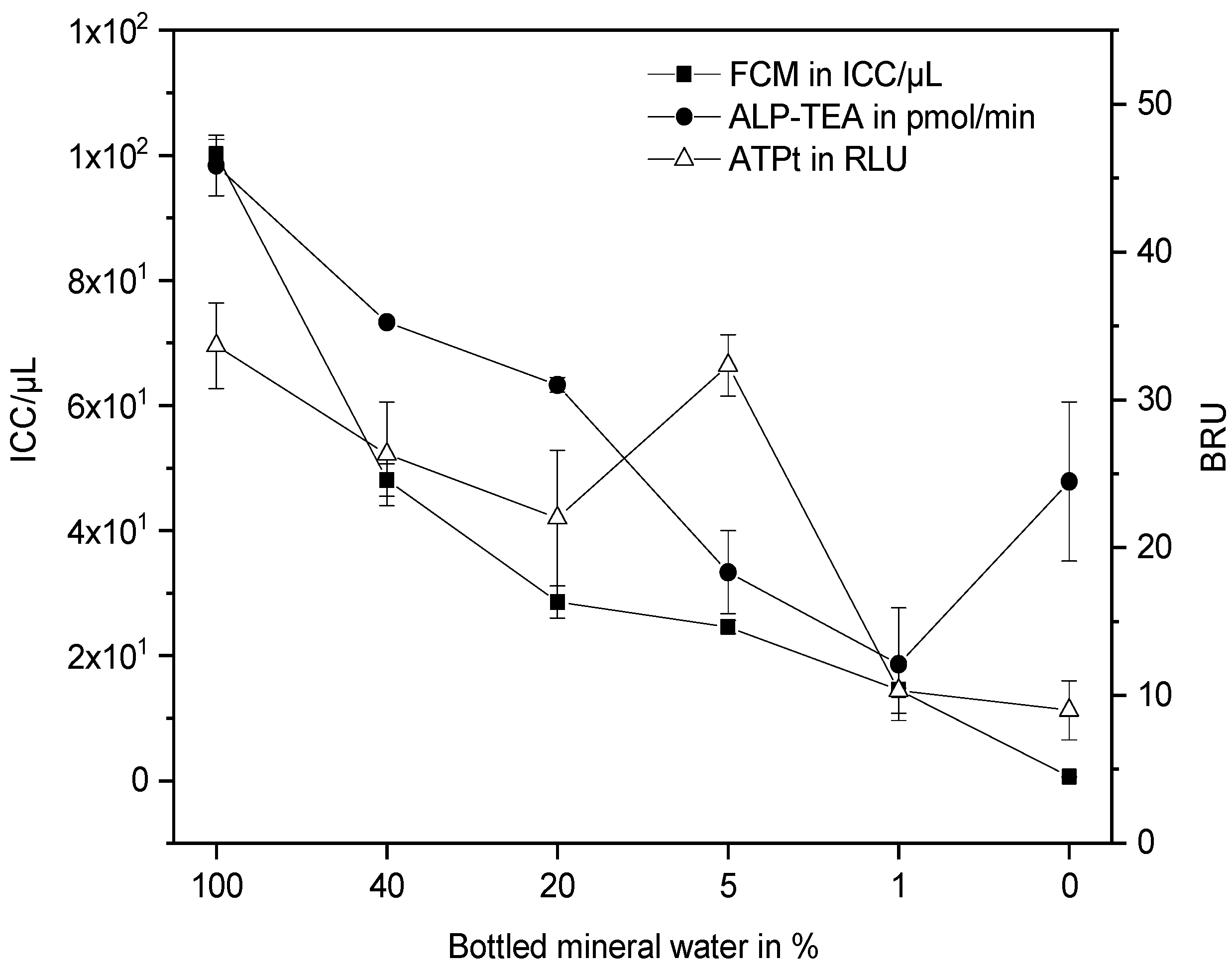
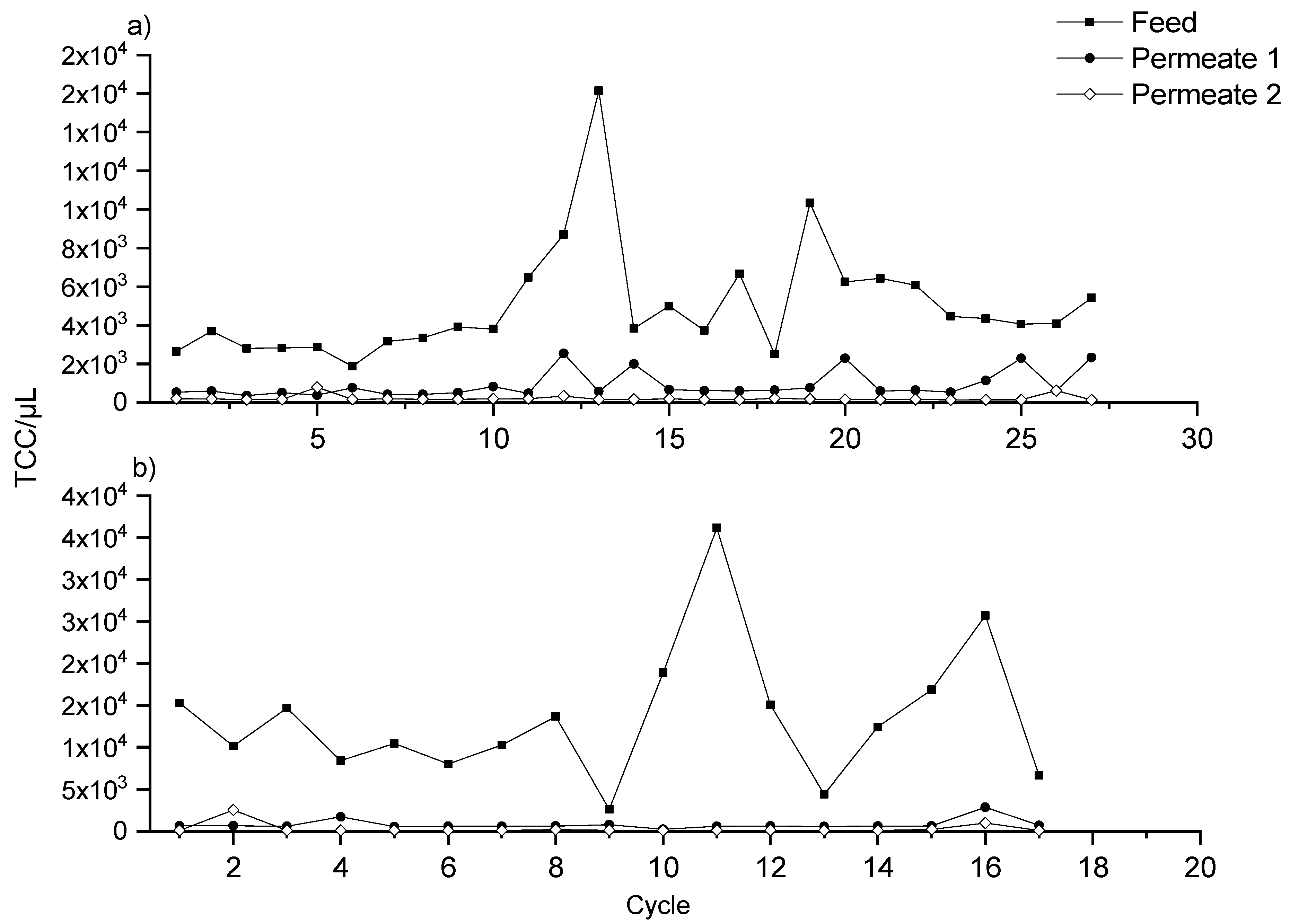
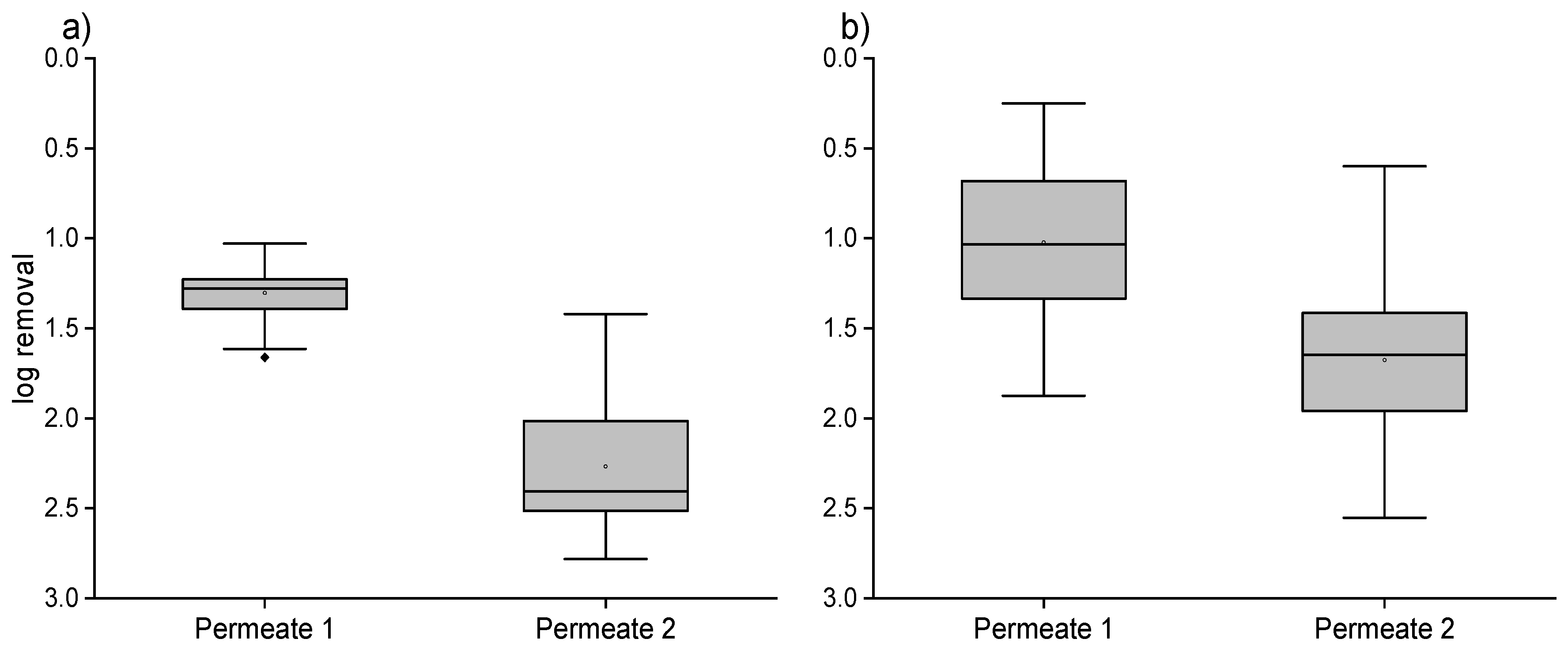
3.2. Ultrafiltration Pilot Plant and Online FCM Measurements
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ALP | Alkaline phosphatase |
| ATP | Adenosine triphosphate |
| MAR | Managed Artificial Recharge |
| BRU | Biomass Related Unit |
| DCC | Desolate Cell Count |
| FCM | Flow cytometry |
| HNA | High Nucleic Acid |
| HNAP | High Nucleic Acid Percentage |
| ICC | Intact Cell Count |
| LNA | Low Nucleic Acid |
| LNAP | Low Nucleoid Acid Percentage |
| RBF | Riverbank filtration |
| TC | Total coliforms |
| TCC | Total Cell Count |
| TEA | Total Enzymatic Activity |
References
- Ray, C.; Jain, R. Drinking Water Treatment: Focusing on Appropriate Technology and Sustainability, 2011th ed.; Springer: Dordrecht, The Netherland; New York, NY, USA, 2011; ISBN 978-94-007-1103-7. [Google Scholar]
- Grischek, T.; Schoenheinz, D.; Worch, E.; Hiscock, K.M. Bank filtration in Europe—An overview of aquifer conditions and hydraulic controls. In Management of Aquifer Recharge for Sustainability; Balkema Publishers, Swets & Zeitlinger: Lisse, The Netherlands, 2002; pp. 485–488. [Google Scholar]
- Ray, C.; Soong, T.W.; Lian, Y.Q.; Roadcap, G.S. Effect of flood-induced chemical load on filtrate quality at bank filtration sites. J. Hydrol. 2002, 266, 235–258. [Google Scholar] [CrossRef]
- De Vera, G.A.; Wert, E.C. Using discrete and online ATP measurements to evaluate regrowth potential following ozonation and (non)biological drinking water treatment. Water Res. 2019, 154, 377–386. [Google Scholar] [CrossRef] [PubMed]
- Allen, M.J.; Edberg, S.C.; Reasoner, D.J. Heterotrophic plate count bacteria—What is their significance in drinking water? Int. J. Food Microbiol. 2004, 92, 265–274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prest, E.I.; Hammes, F.; van Loosdrecht, M.C.M.; Vrouwenvelder, J.S. Biological Stability of Drinking Water: Controlling Factors, Methods, and Challenges. Front. Microbiol. 2016, 7, 45. [Google Scholar] [CrossRef]
- Sun, H.; Shi, B.; Lytle, D.A.; Bai, Y.; Wang, D. Formation and release behavior of iron corrosion products under the influence of bacterial communities in a simulated water distribution system. Environ. Sci. Process Impacts 2014, 16, 576–585. [Google Scholar] [CrossRef]
- Szewzyk, U.; Szewzyk, R.; Manz, W.; Schleifer, K.H. Microbiological safety of drinking water. Annu. Rev. Microbiol. 2000, 54, 81–127. [Google Scholar] [CrossRef]
- World Health Organization. Guidelines for Drinking Water Quality: Incorporating First Addendum; World Health Organization: Geneva, Switzerland, 2006. [Google Scholar]
- Rittmann, B.; Snoeyink, V.L. Achieving biologically stable drinking water. Am. Water Works Assoc. J. 1984, 76, 106–114. [Google Scholar] [CrossRef]
- Bartam, J.; Cotuvo, J.; Exner, M.; Fricker, C.; Glasmacher, A. Heterotrophic Plate Counts and Drinking-Water Safety; IWA Publishing on behalf of the World Health Organization: London, UK, 2003. [Google Scholar]
- Van Nevel, S.; Koetzsch, S.; Proctor, C.R.; Besmer, M.D.; Prest, E.I.; Vrouwenvelder, J.S.; Knezev, A.; Boon, N.; Hammes, F. Flow cytometric bacterial cell counts challenge conventional heterotrophic plate counts for routine microbiological drinking water monitoring. Water Res. 2017, 113, 191–206. [Google Scholar] [CrossRef] [Green Version]
- Hammes, F.; Berney, M.; Wang, Y.; Vital, M.; Köster, O.; Egli, T. Flow-cytometric total bacterial cell counts as a descriptive microbiological parameter for drinking water treatment processes. Water Res. 2008, 42, 269–277. [Google Scholar] [CrossRef]
- Stadler, P.; Loken, L.C.; Crawford, J.T.; Schramm, P.J.; Sorsa, K.; Kuhn, C.; Savio, D.; Striegl, R.G.; Butman, D.; Stanley, E.H.; et al. Spatial patterns of enzymatic activity in large water bodies: Ship-borne measurements of beta-D-glucuronidase activity as a rapid indicator of microbial water quality. Sci. Total Environ. 2019, 651, 1742–1752. [Google Scholar] [CrossRef]
- Højris, B.; Christensen, S.C.B.; Albrechtsen, H.-J.; Smith, C.; Dahlqvist, M. A novel, optical, on-line bacteria sensor for monitoring drinking water quality. Sci. Rep. 2016, 6, 23935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Appels, J.; Baquero, D.; Galofré, B.; Ganzer, M.; van den Dries, J.; Juárez, R.; Puigdomènech, C.; van Lieverloo, J.H. Safety and Quality Control in Drinking Water Systems by Online Monitoring of Enzymatic Activity of Faecal Indicators and Total Bacteria; IWA Publishing: London, UK, 2018; pp. 171–195. ISBN 978-1-78040-869-9. [Google Scholar]
- Vang, Ó.K.; Corfitzen, C.B.; Smith, C.; Albrechtsen, H.-J. Evaluation of ATP measurements to detect microbial ingress by wastewater and surface water in drinking water. Water Res. 2014, 64, 309–320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.J.; Hong, J.T.; Choi, S.J.; Kim, H.S.; Park, W.K.; Han, S.T.; Park, J.Y.; Lee, S.; Kim, D.S.; Ahn, Y.H. Detection of microorganisms using terahertz metamaterials. Sci. Rep. 2014, 4, 4988. [Google Scholar] [CrossRef] [PubMed]
- Haas, R.; Opitz, R.; Grischek, T.; Otter, P. The AquaNES Project: Coupling Riverbank Filtration and Ultrafiltration in Drinking Water Treatment. Water 2019, 11, 18. [Google Scholar] [CrossRef] [Green Version]
- Nagy-Kovács, Z.; László, B.; Simon, E.; Fleit, E. Operational Strategies and Adaptation of RBF Well Construction to Cope with Climate Change Effects at Budapest, Hungary. Water 2018, 10, 1751. [Google Scholar] [CrossRef] [Green Version]
- Vital, M.; Dignum, M.; Magic-Knezev, A.; Ross, P.; Rietveld, L.; Hammes, F. Flow cytometry and adenosine tri-phosphate analysis: Alternative possibilities to evaluate major bacteriological changes in drinking water treatment and distribution systems. Water Res. 2012, 46, 4665–4676. [Google Scholar] [CrossRef] [PubMed]
- Ramseier, M.K.; von Gunten, U.; Freihofer, P.; Hammes, F. Kinetics of membrane damage to high (HNA) and low (LNA) nucleic acid bacterial clusters in drinking water by ozone, chlorine, chlorine dioxide, monochloramine, ferrate(VI), and permanganate. Water Res. 2011, 45, 1490–1500. [Google Scholar] [CrossRef]
- Shirey, T.B.; Thacker, R.W.; Olson, J.B. Composition and stability of bacterial communities associated with granular activated carbon and anthracite filters in a pilot scale municipal drinking water treatment facility. J. Water Health 2012, 10, 244–255. [Google Scholar] [CrossRef] [Green Version]
- Besmer, M.D.; Weissbrodt, D.G.; Kratochvil, B.E.; Sigrist, J.A.; Weyland, M.S.; Hammes, F. The feasibility of automated online flow cytometry for in-situ monitoring of microbial dynamics in aquatic ecosystems. Front. Microbiol. 2014, 5, 265. [Google Scholar] [CrossRef] [Green Version]
- Besmer, M.D.; Epting, J.; Page, R.M.; Sigrist, J.A.; Huggenberger, P.; Hammes, F. Online flow cytometry reveals microbial dynamics influenced by concurrent natural and operational events in groundwater used for drinking water treatment. Sci. Rep. 2016, 6, 38462. [Google Scholar] [CrossRef]
- Egli, T.; Zimmermann, S.; Schärer, P.; Senouillet, J.; Künzi, S.; Köster, O.; Helbing, J.; Montandon, P.-E.; Marguet, J.-F.; Khajehnouri, F. Automatische Online-Überwachung. Bestimmung der Bakterienzahl im Roh- und Trinkwasser: Resultate aus der Praxis. Aqua Gas 2017, 97, 52–59. [Google Scholar]
- Jadoun, J.; Mreny, R.; Saad, O.; Azaizeh, H. Fate of bacterial indicators and Salmonella in biofilm developed on ultrafiltration membranes treating secondary effluents of domestic wastewater. Sci. Rep. 2018, 8, 18066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greenstein, K.E.; Wert, E.C. Using rapid quantification of adenosine triphosphate (ATP) as an indicator for early detection and treatment of cyanobacterial blooms. Water Res. 2019, 154, 171–179. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, M.; Azevedo, N.F.; Ivask, A. Propidium iodide staining underestimates viability of adherent bacterial cells. Sci. Rep. 2019, 9, 6483. [Google Scholar] [CrossRef] [Green Version]
- Kirchhoff, C.; Cypionka, H. Propidium ion enters viable cells with high membrane potential during live-dead staining. J. Microbiol. Methods 2017, 142, 79–82. [Google Scholar] [CrossRef]
- Yang, Y.; Xiang, Y.; Xu, M. From red to green: The propidium iodide-permeable membrane of Shewanella decolorationis S12 is repairable. Sci. Rep. 2015, 5, 18583. [Google Scholar] [CrossRef] [Green Version]
- Nagy-Kovács, Z.; Davidesz, J.; Czihat-Mártonné, K.; Till, G.; Fleit, E.; Grischek, T. Water Quality Changes during Riverbank Filtration in Budapest, Hungary. Water 2019, 11, 302. [Google Scholar] [CrossRef] [Green Version]
- Becker, M.W.; Metge, D.W.; Collins, S.A.; Shapiro, A.M.; Harvey, R.W. Bacterial Transport Experiments in Fractured Crystalline Bedrock. Groundwater 2003, 41, 682–689. [Google Scholar] [CrossRef]
- Ranjan, P.; Prem, M. Schmutzdecke—A Filtration Layer of Slow Sand Filter. Int. J. Curr. Microbiol. Appl. Sci. 2018, 7, 637–645. [Google Scholar] [CrossRef]
- Wang, Y.; Hammes, F.; Boon, N.; Egli, T. Quantification of the Filterability of Freshwater Bacteria through 0.45, 0.22, and 0.1 μm Pore Size Filters and Shape-Dependent Enrichment of Filterable Bacterial Communities. Environ. Sci. Technol. 2007, 41, 7080–7086. [Google Scholar] [CrossRef]
- ReKIS-Viewer. Available online: http://141.30.160.224/fdm/rekisViewer.jsp#menu-5 (accessed on 15 November 2019).
- Vogt, T.; Hoehn, E.; Schneider, P.; Freund, A.; Schirmer, M.; Cirpka, O.A. Fluctuations of electrical conductivity as a natural tracer for bank filtration in a losing stream. Adv. Water Resour. 2010, 33, 1296–1308. [Google Scholar] [CrossRef]
- Hayashi, M.; Vogt, T.; Mächler, L.; Schirmer, M. Diurnal fluctuations of electrical conductivity in a pre-alpine river: Effects of photosynthesis and groundwater exchange. J. Hydrol. 2012, 450–451, 93–104. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time in min | Process | Sample |
|---|---|---|
| 0–5 | backwash | |
| 5-35 | sampling and analysis | Feed |
| 35–40 | backwash | |
| 40–70 | sampling and analysis | Permeate 1 |
| 70–75 | backwash | |
| 75–105 | sampling and analysis | Permeate 2 |
| ATPi | ATPt | HNAP | ICC | TCC | ALP-TEA | |
|---|---|---|---|---|---|---|
| ATPi | 1 | |||||
| ATPt | 1 | |||||
| HNAP | 0.87 | 0.62 | 1 | |||
| ICC | 0.76 | 1 | ||||
| TCC | 0.78 | 1 | ||||
| ALP-TEA | 0.32 | 0.57 | 0.76 | 0.80 | 0.78 | 1 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Adomat, Y.; Orzechowski, G.-H.; Pelger, M.; Haas, R.; Bartak, R.; Nagy-Kovács, Z.Á.; Appels, J.; Grischek, T. New Methods for Microbiological Monitoring at Riverbank Filtration Sites. Water 2020, 12, 584. https://doi.org/10.3390/w12020584
Adomat Y, Orzechowski G-H, Pelger M, Haas R, Bartak R, Nagy-Kovács ZÁ, Appels J, Grischek T. New Methods for Microbiological Monitoring at Riverbank Filtration Sites. Water. 2020; 12(2):584. https://doi.org/10.3390/w12020584
Chicago/Turabian StyleAdomat, Yasmin, Gerit-Hartmut Orzechowski, Marc Pelger, Robert Haas, Rico Bartak, Zsuzsanna Ágnes Nagy-Kovács, Joep Appels, and Thomas Grischek. 2020. "New Methods for Microbiological Monitoring at Riverbank Filtration Sites" Water 12, no. 2: 584. https://doi.org/10.3390/w12020584