Birch Bog on Anthropogenically Transformed Raised Bogs. A Case Study from Pomerania (Poland)
by

Zofia Sotek
1,*, 
Małgorzata Stasińska
1,
Ryszard Malinowski
2,
Renata Gamrat
3 and
Małgorzata Gałczyńska
4 1
Department of Botany and Natural Conservation, Faculty of Biology, University of Szczecin, Felczaka 3c, PL-71-412 Szczecin, Poland
2
Department of Soil Science, Grassland and Environmental Chemistry, West Pomeranian University of Technology Szczecin, Słowackiego 17, PL-71-434 Szczecin, Poland
3
Department of Ecology, Environmental Protection and Management, West Pomeranian University of Technology Szczecin, Słowackiego 17, PL-71-434 Szczecin, Poland
4
Department of Chemistry, Microbiology and Environmental Biotechnology, West Pomeranian University of Technology Szczecin, Słowackiego 17, PL-71-434 Szczecin, Poland
*
Author to whom correspondence should be addressed.
Water 2019, 11(6), 1224; https://doi.org/10.3390/w11061224
Submission received: 1 April 2019
/
Revised: 1 May 2019
/
Accepted: 31 May 2019
/
Published: 12 June 2019
(This article belongs to the Section Hydrology)
Abstract
:Birch bog is formed on the margins of or within raised bogs, on secondary habitats. The study aim was to understand the vegetation and mycological diversity of birch bog on the background of habitat conditions on raised bogs subject to anthropogenic changes, including 15 areas located on seven bogs. Two of the analyzed areas were located on a peat bog not subject to human impact. Phytosociological and mycosociological relevés were taken and substrate analyses were carried out (pH, humidity, N-NH4, N-NO2, N-NO3 and P-PO4). Based on habitat predictors, two area groups were distinguished, differing primarily in humidity. More humid habitats were present on the margins of bogs, and were characterized by lower acidity and higher N-NH4 and P-PO4 abundance. Despite the fact they were enriched by runoffs from the neighboring arable fields, this was not always reflected in the plant and fungi species richness. Quercus robur appeared on less humid habitats, which may be a symptom of unfavorable changes toward habitat drying. In the majority of cases, changes in the habitat independent of the birch patches located and the human impact type are not yet reflected in the vegetation. However, they may be indicated by the fungal diversity, highest in former peat extraction pits, and lowest in pristine peat.
1. Introduction
Bogs are among the most valuable ecosystems, not only due to the rare and valuable species found therein, but also due to the fact that they are natural retention basins. They contain 10% of the freshwater volume of the Earth [1]. Moreover, they are of key importance for the long-term sequestration of atmospheric carbon [2]. It is estimated that peat contains circa 26% of all the terrestrial carbon accumulated since the Last Glacial Maximum [3] thus they are among the largest terrestrial carbon reservoirs. Close to 95% of all peat bogs around the world are located in the northern hemisphere in a cold and at the same time humid temperate climate [4]. It is also here where the majority of raised bogs are present, supplied solely by rainwater. Such ecosystems are highly susceptible not only to natural factors, such as unfavourable climatic changes but above all hydrological disturbances resulting from human activity [5].
Globally, the human impact on wetlands has intensified in the last 200 years, and particularly in the first half of the 20th century [1,6]. This phenomenon occurred at a high rate and extensively primarily in industrialised countries, where the bog areas have been drastically reduced [7]. Wetlands, including raised bogs, were frequently drained in order to use them in agriculture, forestry and to obtain peat [8,9]. In Europe, almost all bog areas have been disturbed [6], whereas in Poland, close to 80% of bog ecosystems have been subject to human interference [10]. In the majority of cases this has led to a loss or degradation of those valuable habitats and, as a consequence, to changes or loss of vegetation typical of bog ecosystems [11,12,13]. Plant species of raised bog communities are typically stenobionts; thus, they are among the ecosystem elements that are most sensitive to habitat changes, including rapidly expanding anthropogenic amendments, including drying and eutrophication [12,14]. Drying may trigger peat decomposition processes, leading to C and CO2 loss, as well as to increased mineralisation of nutrients, leading to internal eutrophication [15,16]. Processes occurring as a result of human impact may lead to a rapid invasion of trees on open areas and development of forest communities. This is an unfavourable phenomenon for raised bogs, as it leads to additional water drainage, increasing their degradation [17,18]. However, in the case of undisturbed bogs, tree invasion is inhibited, and their increasing share takes place only under conditions typical of late stages of natural succession [19].
Among forest communities found on raised bogs, bog coniferous and deciduous forests are distinguished, which are rare and valuable components of those ecosystems. They are included in the Annex 1 to the Habitats Directive of the EU, as 91D0: natural habitats of Community interest whose conservation requires the designation of special areas of conservation [20]. Birch bog (Vaccinio uliginosi-Betuletum pubescentis) is one such community. It is a plant community with Atlantic distribution type, occurring primarily in north-west Europe, including Austria [21], the Czech Republic [22], Denmark [23], Germany [24,25], Hungary [26], Ireland [27], Slovakia [28], and in Ukraine [29]. In Poland, it reaches its eastern boundary of geographic distribution. Patches of birch bog develop typically on drainless terrain depressions with high groundwater level, on rather shallow, mesotrophic, acidic transition peats, on acidic soils with the nature of stagnosol. It is the final stage of succession on transitional bogs, whereas it occurs on the margins of raised bogs, as well as on the habitats with secondary genesis—in the areas where intensive decomposition processes occur [30,31]. In many parts of Ireland, it develops on exploited and drained peatlands, while raised bogs are mainly associated with areas enriched with nutrients, originating from surface runoffs from surrounding areas [32].
Previous studies of swamp birch, both in north-western [23,27,32] and central [21,22] Europe, were mainly focused on understanding the species composition of plants that build this community and its structure. However, in general this aim was not a separate subject of consideration. Usually, it was included in the study on plants colonising the examined bog [33,34,35]; sometimes, the selected properties of the habitat were also taken into account [36]. Few reports concern the Holocene history of birch bog woodlands [37,38,39,40,41]. They show that phases rich in Betula were already present in the early Holocene or the Late Glacial in Central and Eastern Europe.
Apart from plants, fungi are important biotic factors for the formation and functioning of phytocenoses because they are heterotrophic organisms they enter into a range of relationships with plants as well as influence their habitat directly, causing organic matter decomposition. Macromycetes of birch bog are sporadically included in research but they usually are only a single element of wider, more comprehensive studies [42,43]. Certain macromycetes species are strictly associated with peat ecosystems not to be found anywhere else [42,44,45]. Therefore, the condition of the plant community can be reflected not only by plants, their species composition and share, but also the diversity and composition of the mycobiota. These elements remain in a close relationship with the habitat conditions. However, to date these three components were not considered together in the aspect of anthropopression in birch bog. The aim of the study was to investigate the floristic and mycological diversity of birch bog in regard to habitat conditions in raised bogs under anthropogenic transformations. This objective was completed based on the answers to the following research questions: (1) Did the research areas differ significantly in terms of selected physical and chemical properties of the soil, affecting the trophy of the habitat and, if so, whether this was influenced by the type of anthropopression? (2) Are there any significant differences in composition of fungal biota and plant species depending on the location of the research area and the type of anthropogenic impact? (3) If a lower level of habitat predictors (soil properties) is found, is this reflected in both plants and fungi or only in one of these components of the ecosystem? (4) Does the plant community have a chance to preserve its individuality and biodiversity despite the presence of anthropopression?
2. Materials and Methods
The study was conducted in the period 2002–2004 and 2007–2009 in the area of seven raised bogs: Niewiadowo, Roby, Mszar near Stara Dobrzyca, Stramniczka, Torfowisko Toporzyk, Zielone Bagno and Ziemomyśl, located in north-west Poland. Two of the studied objects are mid-forest bogs, and the other ones are located in the close vicinity of agricultural areas, used as arable fields or meadows. This means that they are exposed to an inflow of biogens as a result of surface runoffs. Only Mszar near Stara Dobrzyca was in the past and, is not exposed currently to any anthropogenic influences. Peat exploitation was performed on five of the examined locations, and it was preceded by the dewatering of deposits through a system of ditches that drained water from the peat bogs. However, the time of excavation of the first drainage ditches and the beginning of exploitation is unknown it may be concluded only on the basis of available historical maps. The oldest maps which show the network of drainage ditches and indicate exploitation of the deposits come from 1921–1937 [46,47,48,49]. It suggests, therefore, that peat was being obtained already before this period. The greatest intensity of exploitation probably took place in the first few years after World War II, when peat was the cheapest fuel material. Moreover, the documentation about the moment of finishing the exploitation is lacking. However, reports of the locals show that it was completed in the late 1960s. Currently, peat forming regeneration processes are taking place in the majority of the old excavations, leading to the formation of moss or forest bog phytocenoses. However, dewatering of the peat deposit through the old overgrown ditches continues. At present, with the exception for Ziemomyśl, these bogs are located within Natura 2000 sites and the majority of them are covered by reserves (Table 1).
For the purpose of the study, 15 patches of birch bog (Vaccinio uliginosi-Betuletum pubescentis) representative of the aforementioned bogs were selected (Table 2). Two patches in Mszar near Stara Dobrzyca were treated as reference points for the remaining surfaces due to the fact that they have not been disturbed (no human impact present). The patch sizes were 400 m2, with the exception of patch S2, measuring 200 m2. In order to determine the plant species diversity and their share in the community structure, phytosociological relevés were taken using the Braun-Blanquet method. The patches also constituted permanent plots for the observation of macromycetes. Systematic mycological observations were carried out every 2–3 weeks on average; between 22 and 26 observations were carried out for each plot. The share of fungi in the community was analyzed within bioecological groups: mycorrhizal fungi and saprotrophic fungi (growing on peat and among mosses, litter-inhabiting).
The nomenclature of vascular plants is described in [50] and the nomenclature of mosses is according to [51]. The nomenclature of fungi is given in [52]. The herbarium documentation is deposited in the Herbarium of the Department of Botany and Nature Conservation, Szczecin University (SZUB-F).
Assessment of soil and water conditions was performed based on the analyses of cumulative soil samples. Samples were collected in three consecutive years of the study, on three occasions in the growing season: in spring, summer and autumn (they comprised replications in the statistical analysis), from a depth of 0–20 cm. The following parameters were determined in the samples: ammonium nitrogen content (N-NH4)—via distillation, nitrite nitrogen content (N-NO2)—via distillation, nitrate nitrogen content (N-NO3)—via colorimetry with Griess method, available phosphorus content (P-PO4)—via colorimetry with phosphomolybdate blue (Egner-Riehm method), pH—potentiometry, humidity—by weight in a moisture balance. The analyses were performed at the Department of Soil Sciences, Grassland and Environmental Chemistry of the West Pomeranian University of Technology.
The results concerning the parameters of soils were developed on the basis of the univariate analysis of variance. Additionally, such factors as the place of occurrence of the raised bog, drying up of the habitat, the influence of anthropopressure (surface runoff from agricultural fields and meadows) and the location of the habitat were tested. The significance of differences was evaluated using the Tuckey’s HSD (honestly significant difference) test at the significance level α = 0.05. Based on the standardised physical-chemical data, the investigated soils were grouped by use of hierarchical cluster analysis, Ward’s square Euclidean distance method [53]. This method consists in presenting similarities between objects as a function of distance. The variables describing the object (in our case parameters of soils) are more similar to one another when the distance between them is smaller.
Correlations between plants and fungi and habitat predictors and between plant predictors and fungi were determined by calculating the Pearson’s correlation coefficient. In statistical analyses, the vegetation cover in tree layer ‘a’ and shrub layer ‘b’ was considered jointly, as ‘a + b’, since both layers played a similar role with regards to the group of mycorrhizal fungi and due to the correct interpretation of results they should be analyzed jointly.
These statistical analyses presented in this paper were achieved using the statistical software package for Windows (Statistica® v.12 PL, StatSoft, Szczecin, Poland).
3. Results
The upper layers of peat of all the analyzed areas were characterised by strongly acidic pH and very low, although variable, value of available phosphorus (Table 3). The birch bog in Stramniczka and Zielone Bagno encompasses large areas, forming on dykes and very dry areas, on a layer of humipeat or in poorly hydrated, overgrown former peat extraction pits. In the Stramniczka bog, the upper layers of peat of the S1 and S2 plot have similar humidity and higher amounts of N-NH4 and N-NO2 than N-NO3 (Table 3). On the other hand, plots of the Zielone Bagno bog have variable humidity, yet on ZB2 a significantly higher amount of N-NH4 was found. On Torfowisko Toporzyk, birch bog occurs primarily in the western part of the bog, covered by transition peat deposit, and it remains in contact with alder bog, growing on more eutrophic habitats. In this bog, the upper layer is characterised by high humidity and higher amounts of N-NH4 and N-NO2 than N-NO3. Birch bog on the Niewiadowo bog was formed primarily in its southern and south-western part, creating extensive patches and from the north, it is adjacent to coniferous bog forest. The upper layer of the bog indicates variability in terms of humidity and nitrogen content (at N1 and N2—N-NO3 forms are predominant). The birch bog in Mszar near Stara Dobrzyca covers small areas at the bog margins, indicating a high degree of naturalness. The SD1 and SD2 plots differ significantly in terms of humidity but not soil composition. They have a higher share of the N-NH4 and N-NO2 forms over N-NO3. In the case of the Ziemomyśl, birch bog grows over the majority of its surface, and it is characterised by good hydration of the upper peat layer as well as a high share of the N-NH4 form. On the other hand, at Roby this community occurs primarily on the margins and on slightly dried surfaces (R1-3), which do not exhibit significant variability in terms of humidity and soil chemical composition. The dominance of the N-NO3 form over N-NH4 and N-NO2 is worthy of noting (Table 3).
As a result of habitat predictor analysis based on the estimation of the distance between clusters utilizing analysis of variance (Ward’s method), two major groups of the analyzed plots were obtained (Figure 1). The first group included plots S1, S2, T1, ZB1, ZB2, N1, SD1 and Z1 which, with the exception for T1 plot, were characterized by very high humidity. The second group included plots R1, R2, R3, ZB3, N2, N3 and SD2, which, apart from SD2, were characterized by lower humidity in comparison with the second group.
Both the less humid habitats (on the basis of the division with Ward’s method, Figure 1), as well as human impacts (surface runoffs), indicated significant variability of soils in terms of humidity, pH, ammonium nitrogen and orthophosphate (V) phosphorus. Such differences between soils were not determined for the concentration of nitrate(V) and (III) nitrogen (Table 4 and Table 5). Soils of the study areas exposed to surface runoffs are enriched with N-NH4 and P-PO4 relative to natural and other areas.
Taking into consideration the habitat locality, soils of the analyzed study areas differed statistically significantly only in terms of humidity, soil pH and concentration of ammonium nitrogen (Table 6). The middle parts of the bogs are characterised by higher soil acidity and lower humidity and N-NH4 and P-PO4 abundance than surfaces on the margins and in former peat extraction pits.
Soils of the analyzed study surfaces originating from mid-forest bogs were characterized by significantly higher pHH2O values than those located on mid-field bogs. The remaining physical and chemical parameters of soil were similar (Table 7).
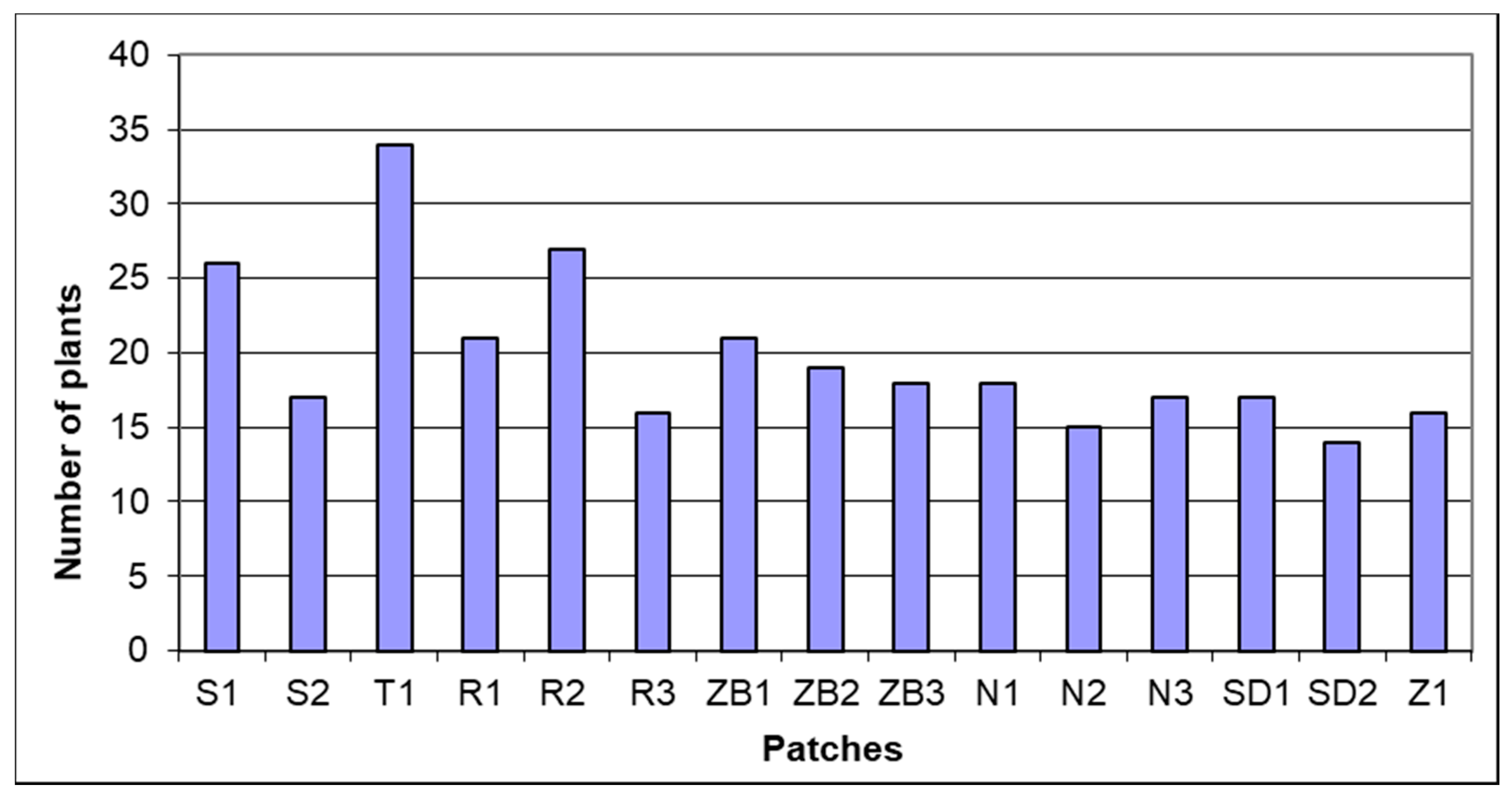
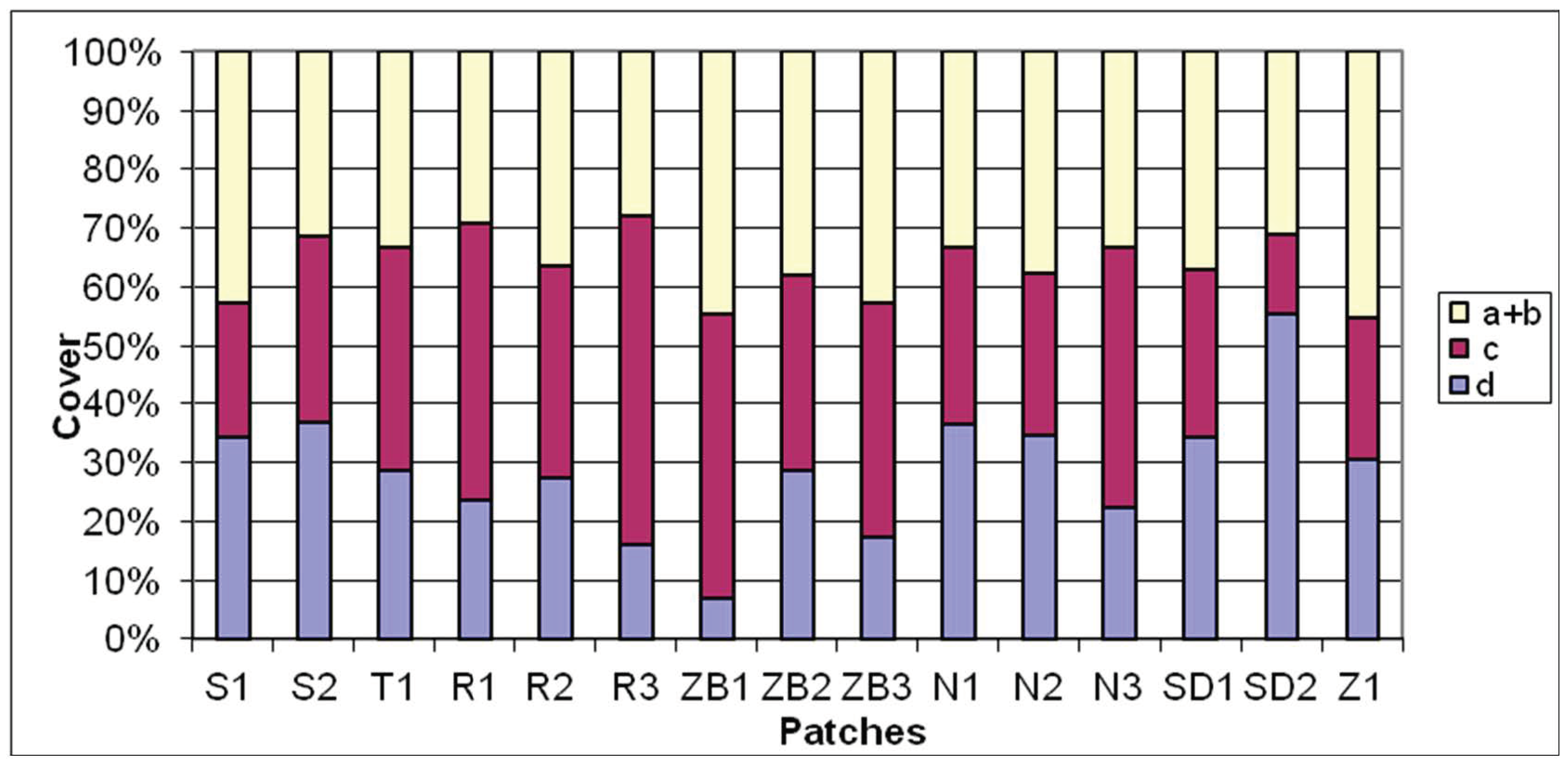
A total of 78 plant taxa were found in birch bog, and in individual patches from 14 to 34 species were recorded (Figure 2). Class Vaccinio-Picetea, in which this community is included, is represented depending on the patch from 2 to 12 species, and moss species from the class Oxycocco-Sphagnetea from 0 to 7. The vegetation cover in certain layers of the examined patches was occasionally variable (Figure 3). This is visible, e.g., in the layers of the tree and shrub stand on Roby bog (R1-3), as compared with the remaining objects. The tree stand of all the analyzed patches is dominated by Betula pubescens, frequently accompanied by Pinus sylvestris (sometimes rather common—N2 and SD1), and less commonly Sorbus acuparia and Betula pendula. The shrub layer is predominantly formed by Betula pubescens and Frangula alnus undergrowth, sometimes with an admixture of Salix spp. In some patches (R1, S2 and SD1-2), the herbal layer is frequented by species of open moss area from class Oxycocco-Sphagnetea, including Erica tetralix, Eriophorum vaginatum and Oxycoccus palustris. The contribution of Sphagnum is marked in the moss layer, and in some areas (SD1-2, S1-2 and T1) abundance of Sphagnum fallax, Sph. palustre and Sph. squarosum, and among brown mosses, Pleurozium schreberi (ZB2 and N1-3) and Aulacomnium palustre (SD2 and R1-2) was recorded. Sphagnum magellanicum was rare, and was recorded only on the preserved habitat (SD1-2). The presence of Lycopodium annotinum—a species regionally characteristic of birch bog was found on T1, N2-3 and ZB1-3. Encroachment of Phragmites australis—a species with wide ecological scale—was recorded in patches R2, S2 and Z1, and a considerable contribution of Molinia cearulea in R1 and R3, with the concomitant appearance of juvenile Quercus robur, whereas Picea abies was growing singularly only in T1.
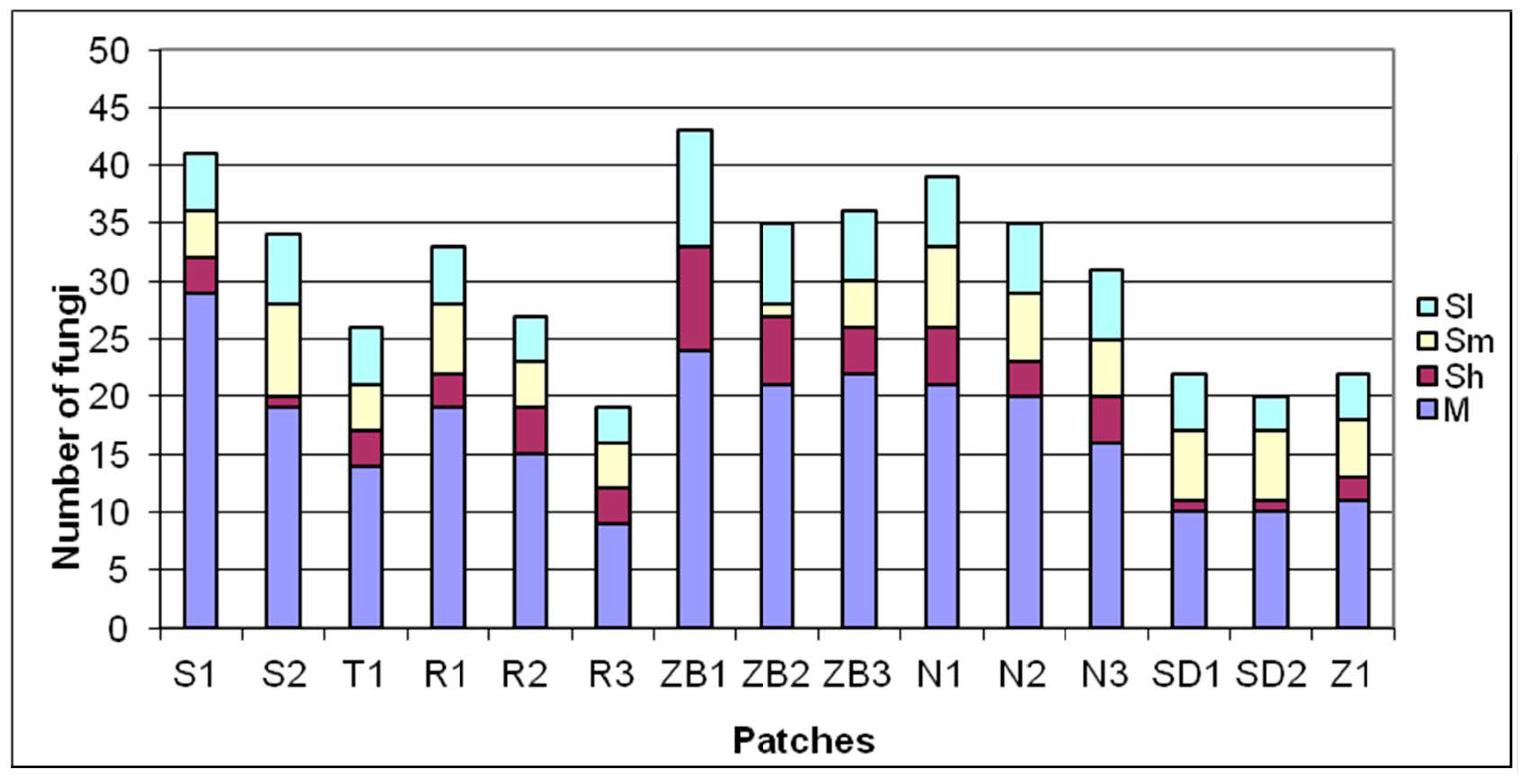
A total of 144 macromycetes species were recorded on the permanent plots of birch bog, and from 33 to 74 species were found on particular plots. As a rule, the highest number of species was found on plots located in former peat extraction pits, and the lowest on bog margins (with the exception of the T1 plot). The share of fungi in the selected ecological groups was variable (Figure 4). Cortinarius flexipes, Laccaria proxima, Lactarius tabidus, Russula betularum and R. claroflava were distinguished among mycorrhizal fungi (46 species) in terms of the frequency of occurrence and abundance. This group was most frequently represented on the plots in former peat extraction pits, and least frequently on bog margins. Lignicolous fungi (47) were predominant among saprotrophic fungi (90), and some of them form permanent, annual or several-year-old fruiting bodies, including Daedaleopsis confragosa, Diatrypella favacea, Fomes fomentarius, Inonotus obliquus and Fomitopsis betulina. The second group of saprotrophic fungi in terms of abundance, including a considerably lower number of species, consisted of peat-growing fungi (15), such as Gymnopus dryophilus, Rhodocollybia maculata, Entoloma cetratum and E. sericatum. Their contribution was variable between individual plots; however, it was lowest on the peat bog margins. Bryophilous fungi (12) were represented by, among others, Bogbodia uda, Hypholoma elongatum and Galerina tibiicystis, and litter-growing fungi (13) by e.g., Mycena galopus and Gymnopus androsaceus. The presence of representatives of both groups on individual plots was variable.
The results of Pearson’s correlation (R) used to determine the type of correlation between plants and fungi and the selected habitat parameters of birch bog indicate the existence of significant relationships between the group of species of class Vaccinio-Picetea, the total number of fungi, M, Sh and Sl fungi groups, and substrate pHKCl, and N-NO3, N-NO2, N-NH4 and P-PO4 content (Table 8). Moreover, the M fungal group exhibited a positive correlation with substrate pHH2O and humidity (R = 0.62, R = 0.64, p < 0.05, respectively). A positive correlation with pHH2O was also found for vegetation cover in ‘a + b’ and ‘d’ layers (R = 0.51, R = 0.56, p < 0.05, respectively). Furthermore, a positive significant relationship occurred between humidity and certain vegetation parameters and the total number of fungi species (Table 9). Moreover, a negative correlation was only found between the species group from class Oxycocco-Sphagnetea, and substrate pHKCl (R = −0.53, p < 0.05).
The existing relationships between fungi and plant predictors are presented in Table 8. A significant positive correlation was obtained between class Vaccinio-Picetea species group, and the total number of fungi species and M, Sh and Sl fungi groups (R = 0.87, R = 0.67, R = 0.72, R = 0.78, p < 0.05, respectively). A significant negative correlation was obtained between class Oxycocco-Sphagnetea species group and the total number of fungi species and Sh fungi group (R = −0.60, R = −0.61, p < 0.05, respectively). Furthermore, the Sh group was negatively correlated with the plant cover of the ‘d’ layer (R = −0.59, p < 0.05, respectively).
4. Discussion
In Poland, birch bog is a rather rare community, restricted to the north-west part of the country. Its mature phytocenoses occupy the habitat with the total area of only 8.75 km2 [30]. Typically, this community is species-poor, which has been observed both in Ireland [27], as well as in Poland, where normally about 20 plant species are found in one patch [30]. Among the birch bog patches we analyzed, 2/3 contained less than 20 plant species (Figure 2), and their low numbers were found not only on the undisturbed peat bog (SD2), but also on certain bog areas subject to human pressure (e.g., N2). Consequently, the low species diversity cannot be linked solely to natural habitats.
Ward’s analysis of birch bog distinguished two habitat groups, less and more humid. The more humid habitats were characterised by lower acidity and were more ammonium nitrogen and phosphorus rich. They were located primarily on the best-hydrated bog margins, which were accessed by runoffs from neighbouring, more nutrient-rich minerotrophic areas. These runoffs often contained biogens from arable lands adjacent to bogs. The patch with the highest floristic diversity (T1, 34 species) was formed on such a habitat. This may suggest that substrate biogen abundance and high humidity play a significant role in both the species richness and composition of the community. However, this was not always reflected by soil richness, for example on Z1—a patch characterised by the highest biogen abundance and high humidity—only 16 plant species were determined on bog margin. Correlation analysis results suggest that the total number of plant species in a community patch is significantly associated only with humidity (Table 8). On the other hand, nutrients such as N-NH4, N-NO3, N-NO2 and P-PO4, may have a positive impact on the abundance of class Vaccinio-Picetea species, in which the discussed community is included. The type of phytocenoses neighboring birch bog significantly influences the number of species found therein. In the case of T1, the neighboring alder bog, rushes and minerotrophic forests resulted in the patch being enriched with plants which penetrated from those habitats, including Peucedanum palustre, Picea abies and Galium palustre. On the other hand, the species poverty of the Z1patch, which was the most biogen-rich, may be associated with i.a. the vicinity of a moss area, which comprises oligotrophic species, exhibiting the lower capacity to penetrate more nutrient-rich habitats. The appearance of Sphagnum magellanicum—a species strictly associated with raised bog habitats, in the SD1-2 patches, located on bog margins and not subject to human pressure, also resulted from the direct vicinity of moss area with birch bog. The negative correlation between the number of species from class Oxycocco-Sphagnetea, and pHKCl (Table 8) is typically explained by the considerable share of Sphagnum spp., as they additionally impact habitat acidification, by binding cations (Ca and Mg) present in the environment and the release of hydrogen ions [54]. However, at Roby (lowest pH) this correlation can be linked with slightly higher (with regards to the majority of the remaining study objects) number of herbaceous plant species included in this class, and not with Sphagnum spp., as those mosses grew there only at low amounts. Typically, the share of raised bog species in birch bog is low [30].
Less humid habitats were mostly located within the bogs. The least humid birch bog patches were located in Roby (R1-3) and Niewiadów (N2). They were richer in N-NO3 than N-NH4, which could be a symptom of unfavourable changes toward habitat drying [55,56,57]. This phenomenon may be also suggested by the presence of single juvenile Quercus robur individuals [58] recorded in patches R1 and R3, where moor grass was also found, as well as in R2. Moreover, in the vegetation structure of R2, a high share of shrub ‘b’ layer was recorded, sometimes higher than trees ‘a’, this could have stemmed from the lower habitat humidity, which may have promoted numerous appearances of Betula pubescens in the understory which was definitely predominant here.
The spatial structure of birch bog was in some cases otherwise, e.g., by comparing herbs layer ‘c’ and moss layer ‘d’ on extracted bogs with undisturbed bog (Mszar near Stara Dobrzyca). In bog patches subject to human change, the ‘c’ cover was usually higher than the ‘d’. A reverse relationships were found in Betuletum pubescentis on the previously exploited peatlands in Ireland [27], as well as in a similar community described as bogged birch forest with peat located on similar disturbed habitats in Siberia [42]. Former peat extraction pits, in which birch bog (S1-2, ZB1-3, N1 and N3) was formed, were characterised by similar humidity and slightly lower acidity in comparison with areas on the bog margins. However, the vegetation did not demonstrate considerable differences with relation to this community patches located on other sites. In these sites, birch bog was typically well-developed, which was also indicated by the presence of Lycopodium annotinum in half of these patches, a characteristic species for this community, as well as common Pleurozium schreberi, which had a considerable share in some patches. Former peat extraction pits constitute secondary habitat for birch bog and it may thrive in them only when the high water level is not maintained in this habitat, otherwise Betula pubescens will gradually wither, and under favorable conditions succession may have the direction of moss community restoration. Expansion of forest vegetation is determined by the groundwater level in the bog and the air availability in the upper peat layer [57,59]. For the normal development of the root system, trees require more than a 10% share of peat pores to be filled with air [59]. The analyzed bogs, independently of the study area location (middle, margin, former peat extraction pits), fulfilled this condition, only on peat bog not subject to human impact, the study area SD1 had pores filled with air in 8% and with water in 92%. Furthermore, trees require a nutrient supply for normal development. Birch requires considerably higher amounts of nutrients than pine [60]. On no disturbed peat bogs has the development of trees been inhibited primarily by the excessive humidity and bog growth, and to a lesser degree by the nutrient deficiency [61,62].
Molinia cearulea is a common element of birch bog and its share in this community may vary. In Austria, apart from Vaccinium uliginosum, it is a common predominant plant species of the herbs layer of this community [21]. In patches R1 and R3, where moor grass was most common, it was observed that peat mosses were characterised by lower cover percentage. Competition between these plants for light could be one of the causes of this phenomenon. The importance of this factor was indicated by Hogg et al. [63], who observed that cutting back M. coerulea restricts its competitiveness for light, having a positive impact on the development of peat mosses. On the other hand, Limmper [64] determined that the high share of herbaceous plants on peat bogs may have a negative impact on the development of these mosses. The shadow effect by vascular plants has been indicated to be a cause for this state by Hayward and Clymo [65] and by Heijmans et al. [66]. In turn, the living layer of peat moss may inhibit the growth of vascular plants by binding large amounts of nitrogen [64,67].
Birch and moor grass may appear on ombrotrophic bogs not only on their natural habitats, that is the margins of raised bog, but also on moss areas. It is a highly unfavourable phenomenon as the increase of their total cover and expansion of other vascular plants stimulates evapotranspiration [67,68]. This may contribute to habitat drying, which is very sensitive to changes in water relations, and in such cases susceptible to invasion of Picea abies, which is an alien species in terms of habitat and geography. However, no such phenomenon has been observed on the analyzed patches located in the middle of the bogs. The presence of singular individuals of spruce was recorded only in T1, located in a more nutrient-rich bog margin.
Birch bog is one of the bog forest communities least known in mycological terms. Fungi constitute its important, integral structural element, as they enter into a series of interactions with different plant species and they influence not only the maintenance but also the habitat transformation [69]. They also play the dominant role among decomposer organisms in acidic peat ecosystems [44]. The majority of them are saprotrophs, participating in the decomposition of organic matter [70,71]. Birch bog, other than for plants, is typically rich in macroscopic fungi species, which has been determined in both the Słowiński National Park (78 species, [72]) as well as in the Goleniów Forest (85 species, [73]). Among the analyzed plots, ZB1-2 and N1 (61-74 species) are the richest in fungi species, and are located in former peat extraction pits. In turn, the SD1-2 plots not subject to human pressure turned out to be the least rich in fungi species (33–39). The contribution of peat mosses could be one of the factors restricting the number of fungi species on an undisturbed habitat, as it was here highest among all the analyzed plots. The locally compact layer of peat mosses typically inhibited the development of fungi growing on a different substrate, including peat, which is further suggested by the negative correlation, whereas it favored the occurrence of a small group of fungi associated with moss areas, e.g., Galerina paludosa, G. tibiicystis and Sphagnurus paluster. The role of these moss species is confirmed by the obtained negative correlation between the total number of fungi species per plot, and the number of plant species from class Oxycocco-Sphagnetea. An analysis of Pearson’s correlation indicates that both the total number of species of fungi found within one surface and the number of mycorrhoidal species were significantly and positively correlated with the number of plant species from the Vaccinio-Picetea class. This relationship stems from the community structure in which the Vaccinio-Picetea class plant species are among its basic elements, and in particular B. pubescens and its accompanying trees interacting with fungi. Numerous fungi species, including mycorrhizal species, are specific for given tree species [74], which is reflected by the biota of macromycetes of the given community. Plots N2 (in the middle of the bog) and SD1 (bog margin) are examples here, where the considerable share of pine resulted in the appearance of fungi associated with them, including Russula emetica, Lactarius rufus and Auriscalpium vulgare.
The fungal species diversity of birch bog does not only depend on the multi-plane relationships between these specific organisms and plants but also on chemical properties of the soil. Results presented in this study indicate the existence of significant relationships between the occurrence of fungi species and selected environmental factors, i.a. humidity and soil pH. In birch bog, all the study areas were characterised by more or less acidic soil. The determined positive correlation between the number of fungi and certain bioecological fungi groups, and pH suggests that excessively acidic soil may have a negative impact on the diversity of macromycetes. Numerous fungi species prefer determined values of pH and react differently to its change [75,76]. A similar situation occurs in the case of phosphorus and nitrogen compound content in soil, as many fungi species e.g., ectomycorrhizal, exhibit different tolerance to the high or low content of nitrogen in soil [77]. Moreover, the availability of different nitrogen forms plays a significant role in the production of fruiting bodies [76] and formation of ectomycorrhiza [78]. Based on the obtained positive correlation between the total number of macromycetes on the given plot and certain bioecological groups of fungi, as well as the content of phosphorus and various nitrogen forms in the soil it can be expected that with their increase macromycetes diversity will increase. Trees growing on humid peat soils largely depend on ectomycorrhizal symbionts which facilitate their absorption of nutrients and participate in capturing P [79,80]. In the case of Betula pubescens saplings [67], it was determined that the presence of ectomycorrhizal symbionts did not guarantee trees growth, which could stem from inhibition of the activity of those symbionts by acidic conditions. The correct content of nitrogen and phosphorus in the soil is of particular importance for the development of birch [11,67], given that birch is incapable of fully utilising nitrogen in the event of the absence of P in soil [81].
5. Conclusions
Changes occurring in the habitat of Vaccinio uliginosi-Batuletum pubescentis birch bog regardless of (1) location of bog patches, (2) anthropopression type (drainage, peat exploitation, surface runoff), (3) time of anthropogenic impact (peat exploitation - lasted about 40 years, completed approx. 60 years ago, dewatering—started a few years before exploitation of peat, lasts with lower intensity to today) and (4) intensity of human impact, in most cases are not yet reflected in the vegetation, as found on the basis of a comparison with birch growing in a peat bog not disturbed by humans. Plant species, e.g., Quercus robur, started to appear only in singular areas, but in the case of their greater share they may have indicate ongoing transformations of these habitats. Habitat conditions enable the presence of only a strictly limited number of widely distributed plant species, such as Phragmites austrialis. However, the slow rate of change is demonstrated by the high diversity of fungi species, which was highest in the areas of former peat extraction pits, whereas lowest on bog margins, and particularly on the undisturbed bog. On the other hand, the considerable presence of fungi that are strictly associated with bogs constitutes an indicator for the still good preservation state of the studied objects.
When birch bog appears on secondary habitats in the middle of a bog, a dilemma arises as to whether humans should allow for its further development. In the case where birch bog would threaten the existence of a moss area on an ombrogenic peat bog, activities aiming toward its eradication for the advantage of the moss area should be undertaken by using e.g., long-term submerging of the area where birch bog is found. However, if this community is well developed, which would be demonstrated by, i.a. presence of its characteristic species, and its state, based on the accepted indicators, can be evaluated as good [58], then such a community should be protected also on secondary habitats, as it is rare and has priority among Natura 2000 habitats. It is a permanent community in stable hydrological conditions. Despite the fact that the majority of the studied birch bog habitats have been and still are being mostly subject to different types of human impact, their physicochemical properties, including humidity, enable further development of this community. The majority of the analyzed birch bog patches are located on protected areas, thus granting them a higher chance of survival.
Author Contributions
Z.S. and M.S. designed, conducted the research, partly analyzed the data, and wrote the paper; R.M. carried out laboratory analyzes, partly analyzed the data and participated in writing; R.G. participated in fieldwork and reviewed the final draft of the manuscript; M.G. partly analyzed the data and reviewed the final draft of the manuscript. All authors read and approved the final manuscript.
Funding
Studies supported financially in part by the Ministry of Science and Higher Education (Poland), grant N N305 2617 33.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Joosten, H.; Clarke, D. Wise Use of Mires and Peatlands; International Mire Conservation Group and International Peat Society: Saarijarvi, Finland, 2002. [Google Scholar]
- Dise, N.B.; Narasinha, J.S.; Weishampel, P.; Verma, S.B.; Verry, E.S.; Gorham, E.; Crill, P.M.; Harriss, R.C.; Kelley, C.A.; Yavitt, J.B.; et al. Carbon emissions from peatlands. In Peatland Biogeochemistry and Watershed Hydrology at the Marcell Experimental Forest; Kolka, R.K., Sebestyen, S.D., Verry, E.S., Brooks, K.N., Eds.; CRC Press Taylor & Francis Group: Boca Raton, FL, USA; London, UK; New York, NY, USA, 2011; pp. 297–347. [Google Scholar]
- Smith, L.C.; MacDonald, G.M.; Velichko, A.A.; Beilman, D.W.; Borisova, O.K.; Frey, K.E.; Kremenetski, K.V.; Sheng, Y. Siberian Peatlands a Net Carbon Sink and Global Methane Source Since Early Holocene. Science 2004, 303, 353–356. Available online: http://science.sciencemag.org/content/sci/303/5656/353 (accessed on 5 February 2019). [CrossRef] [PubMed]
- Franzén, L.G. Increased Decomposition of Subsurface Peat in Swedish Raised Bogs: Are Temperate Peatlands Still Net Sinks of Carbon? Mires Peat 2006, 1, 3. Available online: http://pixelrauschen.de/wbmp/media/map01/map_1_3.pdf accessed (accessed on 10 February 2019).
- Andersen, R.; Rochefort, L.; Poulin, M. Peat, Water and Plant Tissue Chemistry Monitoring: A Seven-Year Case-Study in a Restored Peatland. Wetlands 2010, 30, 159–170. Available online: https://link.springer.com/article/10.1007/s13157-009-0015-0 (accessed on 8 February 2019). [CrossRef]
- Minayeva, T.; Sirin, A.; Bragg, O. (Eds.) A Quick Scan of Peatlands in Central and Eastern Europe; Wetlands International: Wageningen, The Netherlands, 2009; 132p. [Google Scholar]
- Franzén, L.G.; Chen, D.; Klinger, L.F. Principles for a Climate Regulation Mechanism during the Late Phanerozoic era, Based on Carbon Fixation in Peat-Forming Wetlands. Ambio 1996, 25, 435–442. Available online: https://inis.iaea.org/search/search.aspx?orig_q=RN:28025142 (accessed on 9 February 2019).
- Strack, M. (Ed.) Peatlands and Climate Change; IPCC: Saarijärvi, Finland, 2008. [Google Scholar]
- Turunen, J. Development of FINNISH Peatland Area and Carbon Storage 1950–2000. Boreal Environ. Res. 2008, 13, 319–334. Available online: https://helda.helsinki.fi/bitstream/handle/10138/234741/ber13-4-319.pdf?sequence=1 (accessed on 9 February 2019).
- Sienkiewicz, J.; Kloss, M. Distribution and conservation of mires in Poland. In Regional Variation and Conservation of Mire Ecosystems; Moen, A., Ed.; NTNU University Museum, Gunneria: Trondheim, Norway, 1995; Volume 70, pp. 149–158. [Google Scholar]
- Tomassen, H.B.M.; Smolders, A.J.P.; Limpes, J.; Lamers, L.P.M.; Roelofs, J.G.M. Expansion of invasive species on ombrotrophic bogs: Desiccation or high N deposition? J. Appl. Ecol. 2004, 41, 139–150. [Google Scholar] [CrossRef]
- Kollmann, J.C.; Rasmussen, K.K. Succession of a degraded bog in NE Denmark over 164 years–monitoring one of the earliest restoration experiments. Tuexenia 2012, 32, 67–85. [Google Scholar]
- Sotek, Z. Distribution Paterns, History, and Dynamics of Peatland Vascular Plants in Pomerania (NW Poland). Biodiv. Res. Conserv. 2010, 18, 1–82. Available online: https://www.degruyter.com/downloadpdf/j/biorc.2010.18.issue--1/v10119-010-0020-4/v10119-010-0020-4.pdf (accessed on 8 February 2019).
- Herbichowa, M.; Pawlaczyk, P.; Stańko, R. Conservation of Baltic Raised Bogs in Pomerania. In Experience and Results of the LIFE04NAT/PL/000208 PLBALTBOGS Project; Wydawnictwo Klubu Przyrodników: Świebodzin, Poland, 2007. (In Polish) [Google Scholar]
- Smolders, A.J.P.; Lamers, L.P.M.; Lucassen, E.C.H.E.T.; Van der Velde, G.; Roelofs, J.G.M. Internal eutrophication: ‘how it works and what to do about it’, a review. Chem. Ecol. 2006, 22, 93–111. [Google Scholar] [CrossRef]
- Lamers, L.P.; Vile, M.A.; Grootjans, A.P.; Acreman, M.C.; van Diggelen, R.; Evans, M.G.; Richardson, C.J.; Rochefort, L.; Kooijman, A.M.; Roelofs, J.G. Ecological Restoration of Rich Fens in Europe and North America: From Trial and Error to an Evidence-Based Approach. Biol. Rev. 2014, 90, 182–203. Available online: https://onlinelibrary.wiley.com/doi/full/10.1111/brv.12102 (accessed on 8 February 2019). [CrossRef]
- Fay, E.; Lavoie, C. The Impact of Birch Seedlings on Evapotranspiration from a Mined Peatland: An Experimental Study in Southern Quebec, Canada. Mires Peat 2009, 5, 1–7. Available online: http://citeseerx.ist.psu.edu/viewdoc/download?doi=10.1.1.493.7184&rep=rep1&type=pdf (accessed on 8 February 2019).
- Talbot, J.; Richard, P.J.H.; Roulet, N.T.; Booth, R. KAssessing long-term hydrological and ecological responses to drainage in a raised bog using paleoecology and a hydrosequence. J. Veg. Sci. 2010, 21, 143–156. [Google Scholar] [CrossRef]
- Gunnarsson, U.; Malmer, N.; Rydin, H. Dynamics or constancy in Sphagnum dominated mire ecosystems? A 40-year study. Ecography 2002, 25, 685–704. [Google Scholar] [CrossRef] [Green Version]
- Directive, H. Directive 92/43/EEC of 21 May 1992 on the conservation of natural habitats and of wild fauna and flora. Off. J. Eur. Union 1992, 206, 7–15. [Google Scholar]
- Mucina, L.; Grabherr, G.; Wallnöver, S. Die Pflanzengesellschaften Österreichs; Wälder und Gebüsche. Gustav Fischer: Jena, Germany, 1993; Volume 3, pp. 1–350. [Google Scholar]
- Horodyská, E. Minerotrofní rašeliniště Českomoravské vrchoviny: Srovnání historických a současných dat; Ms., dipl. pr., PřF MU: Brno, Czech Republic, 2006. [Google Scholar]
- Dierßen, K. Vegetation Nordeuropas; Verlag Eugen Ulmer: Stuttgart, Germany, 1996. [Google Scholar]
- Oberdorfer, E. Süddeutsche Pflanzengesellschaften; Wälder und Gebüsche; Gustav Fischer: Jena, Germany, 1992; Volume 4, pp. 1–862. [Google Scholar]
- Preising, E.; Weber, H.E.; Vahle, H.-C. Die Pflanzengesellschaften Niedersachsens—Wälder und Gebüsche. Naturschutz Landschaftspflege Niedersachsen; Nds. Landesbetrieb für Wasserwirtschaft, Küsten- und Naturschutz: Hannover, Germany, 2003; Volume 20, pp. 1–139. [Google Scholar]
- Kevey, B. Magyarország erdötársulásai. Tilia 2008, 14, 1–489. Available online: http://real-d.mtak.hu/152/1/Kevey.pdf (accessed on 5 February 2019).
- Kelly, D.L.; Iremonger, S.F. Irish wetland woods: The plant communities and their ecology. In Biology and Environment: Proceedings of the Royal Irish Academy; Royal Irish Academy: Dublin, Ireland, 1997; Volume 97B, pp. 1–32. Available online: https://www.jstor.org/stable/20499982 (accessed on 7 February 2019).
- Michalko, J.; Berta, J.; Magic, D. Geobotanická mapa ČSSR; Slovenská socialistická republika: Veda, Bratislava, 1986. [Google Scholar]
- Didukh, Y.P.; Fitsailo, T.V.; Korotchenko, I.A.; Iakushenko, D.M.; Pashkevych, N.A. Biotopy lisovoï ta lïsostepovoï zon Ukraïny; TOV MAKROS: Kyïv, Ukraine, 2011. [Google Scholar]
- Herbichowa, M. Bory i lasy bagienne. B. Opis podtypów. Brzezina bagienna. In Lasy i bory. Poradnik ochrony siedlisk i gatunków Natura 2000—Podręcznik metodyczny; Herbich, J., Ed.; Ministerstwo Środowiska: Warszawa, Poland, 2004; Volume 5, pp. 175–178. (In Polish) [Google Scholar]
- Matuszkiewicz, W. A guide for marking Poland’s plant communities. In Biodiversity of Poland; Mirek, Z., Ed.; Inst. Bot. im. W. Szafera. PAN: Kraków, Poland, 2002; Volume 3, p. 537. (In Polish) [Google Scholar]
- Cross, J.R. Unusual stands of birch on bogs. Irish Nat. J. 1987, 22, 305–310. Available online: https://www.jstor.org/stable/25539176?seq=1#page_scan_tab_contents (accessed on 25 April 2019).
- Brzeg, A.; Kuświk, H.; Melosik, I.; Urbański, P. Flora and vegetation of the projected nature reserve “Torfowisko Toporzyk” in Drawski Landscape Park. Bad. Fizjogr. Pol. Zach. B 1995, 44, 51–76. (In Polish) [Google Scholar]
- Brzeg, A.; Kuświk, H.; Melosik, I.; Urbański, P. Flora and vegetation of the projected nature reserve “Zielone Bagna” in Drawski Landscape Park. Bad. Fizjogr. Pol. Zach. B 1996, 45, 121–145. (In Polish) [Google Scholar]
- Sotek, Z.; Stasińska, M. Diversity of macromycetes and the transformation of vegetation in the atlantic raised bog, Stramniczka”. Woda-Środowisko-Obszary Wiejskie 2010, 10, 257–270. (In Polish) [Google Scholar]
- Sotek, Z.; Grzejszczak, G.; Stasińska, M.; Malinowski, R. Synanthropisation of the Baltic-type raised bog “Roby” (NW Poland). Biodiv. Res. Conserv. 2015, 38, 51–56. [Google Scholar]
- Bešta, T.; Šafránková, J.; Pouzar, M.; Novák, J.; Nováková, K. Late Pleistocene—Early Holocene transition recorded in the sediments of a former shallow lake in the Czech Republic. Hydrobiologia 2009, 631, 107–120. [Google Scholar] [CrossRef]
- Gálová, A.; Hájková, P.; Petr, L.; Hájek, M.; Jamrichova, E. Origin of a boreal birch bog woodland and landscape development on a warm low mountain summit at the Carpathian-Pannonia interface. Holocene 2016, 26, 1112–1125. [Google Scholar] [CrossRef]
- Karpińska-Kołaczek, M.; Kołaczek, P.; Stachowicz-Rybka, R. Palaeobotanical studies on Late Glacial and Holocene vegetation development and transformations of the ‘Wielkie Błoto’ mire near Gołdap (north-eastern Poland). Acta Palaeobot. 2013, 53, 53–67. [Google Scholar] [CrossRef]
- Karpińska-Kołaczek, M.; Kołaczek, P.; Stachowicz-Rybka, R. Pathways of woodland succession under low human impact during the last 13,000 years in northeastern Poland. Quat. Int. 2014, 328–329, 196–212. [Google Scholar] [CrossRef]
- Kołaczek, P.; Gałka, M.; Karpińska-Kołaczek, M. Succession of arboreal taxa during the Late Glacial in south-eastern Poland: Climatic implications. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2015, 421, 1–14. [Google Scholar] [CrossRef]
- Filippova, N.V.; Bulyonkova, T.M. The diversity of larger fungi in the vicinities of Khanty-Mansiysk (middle taiga of West Siberia). Environ. Dyn. Glob. Clim. Chang. 2017, 8, 13–24. Available online: http://journals.eco-vector.com/EDGCC/article/view/6949 (accessed on 11 February 2019). [CrossRef]
- Stasińska, M. Macrofungi of raised and transitional bogs of Pomerania. Monogr. Bot. 2011, 101, 1–142. [Google Scholar] [CrossRef]
- Thormann, M.N.; Rice, A.V. Fungi from Peatlands. Fungal Divers. 2007, 24, 241–299. Available online: http://www.cfs.nrcan.gc.ca/bookstore_pdfs/29484.pdf (accessed on 9 February 2019).
- Chinan, V. Macrofungi Identified in the Peat Bogs from the NATURA 2000 Site “Găina Lucina” (Eastern Carpathians, Romania). Analele ştiinţifice ale Universităţii “Al. I. Cuza” Iaşi. Biologie Vegetală 2010, 56, 61–64. Available online: http://www.bio.uaic.ro/publicatii/anale_vegetala/issue/2010F1/08-2010F1.pdf (accessed on 8 February 2019).
- Mapa topograficzna. 158. Czaplinek (Tempelburg) 1:100 000; Instytut Wojskowo-Geograficzny: Warszawa, Poland, 1921; Available online: http://maps.mapywig.org/m/WIG_maps/series/100K/158_CZAPLINEK_(Tempelburg)_1921_LoC_G6520_s100_.P6.jpg (accessed on 24 April 2019).
- Mapa topograficzna. 92. Treptow a. d. Rega (Trzebiatowo) 1:100 000. (Druk na podstawie mapy 1:100 000 wydania z r. 1929); Wojskowy Instytut Geograficzny: Warszawa, Poland, 1937; Available online: http://maps.mapywig.org/m/WIG_maps/series/100K_300dpi/P32_S20_92_TREPTOW_a.d._REGA_(TRZEBIATOWO)_1937_300dpi.jpg (accessed on 24 April 2019).
- Mapa topograficzna. 93. Kolberg (Kołobrzeg) 1:100 000. (Druk na podstawie mapy 1:100 000 wydania z r. 1929. Częściowo uzupełniono); Wojskowy Instytut Geograficzny: Warszawa, Poland, 1937; Available online: http://www.ziemiagryfa.org.pl/wp-content/uploads/2014/12/P32_S21_93_KOLBERG_KOLOBRZEG_1937_300dpi.jpg (accessed on 24 April 2019).
- Mapa topograficzna. 156. Naugard (Nowogard) 1:100 000. (Druk na podstawie mapy 1:100 000 wydania z r. 1929. Częściowo uzupełniono); Wojskowy Instytut Geograficzny: Warszawa, Poland, 1937; Available online: http://maps.mapywig.org/m/WIG_maps/series/100K_300dpi/P34_S20_156_NAUGARD_(NOWOGARD)_1936_300dpi.jpg (accessed on 24 April 2019).
- Mirek, Z.; Piękoś-Mirkowa, H.; Zając, A.; Zając, M. Flowering plants and Pteridophytes of Poland—A checklist. In Biodiversity of Poland; Mirek, Z., Ed.; Inst. Bot. im. W. Szafera. PAN: Kraków, Poland, 2002; Volume 1, p. 442. [Google Scholar]
- Ochyra, R.; Żarnowiec, J.; Bednarek-Ochyra, H. Census catalogue of Polish mosses. In Biodiversity of Poland; Mirek, Z., Ed.; Inst. Bot. im. W. Szafera. PAN: Kraków, Poland, 2003; Volume 3, p. 372. [Google Scholar]
- Index Fungorum. Database 2016. Available online: http://www.indexfungorum.org/Names/Names.asp (accessed on 15 February 2019).
- Hair, J.F.; Black, W.C.; Babin, B.J.; Anderson, R.E. Multivariate Data Analysis, 7th ed.; Pearson Education Limited: London, UK, 2014. [Google Scholar]
- Pawlaczyk, P.; Herbichowa, M.; Stańko, R. Protection of the Baltic Bogs: A Guide for Practitioners, Theorists and Officials; Wydawnictwo Klubu Przyrodnikow: Świebodzin, Poland, 2005. (In Polish) [Google Scholar]
- Salonen, V. Revegetation of harvested peat surfaces in relation to substrate quality. J. Veg. Sci. 1994, 5, 403–408. [Google Scholar] [CrossRef]
- Wind-Mulder, H.L.; Rochefort, L.; Vitt, D.H. Water and peat chemistry cutover of natural and post-harvested peatlands across Canada and their relevance to peatland restoration. Ecol. Eng. 1996, 7, 161–181. [Google Scholar] [CrossRef]
- Zając, E.; Zarzycki, J.; Ryczek, M. Substrate Quality and Spontaneous Revegetationof Extracted Peatland: Case Study of an Abandoned Polish Mountain Bog. Mires Peat 2018, 21, 1–14. Available online: https://www.researchgate.net/profile/Ewelina_Zajac/publication/326172848_Degradation_of_peat_surface_on_an_abandoned_post-extracted_bog_and_implications_for_re-vegetation/links/5b4c5d12a6fdccadaecf5c46/Degradation-of-peat-surface-on-an-abandoned-post-extracted-bog-and-implications-for-re-vegetation.pdf (accessed on 3 March 2019).
- Pawlaczyk, P. Bory i lasy bagienne. In Monitoring siedlisk przyrodniczych. Przewodnik metodyczny; Mróz, W., Ed.; Biblioteka Monitoringu Środowiska: Warszawa, Poland, 2010; Volume 1, pp. 216–235. [Google Scholar]
- Paavilainen, E.; Pälvänen, J. Peatland Foresty: Ecology and Principles; Ecological Studies 111; Springer: Berlin, Germany, 1995; pp. 1–256. [Google Scholar]
- Wieder, R.K.; Vitt, D.H. (Eds.) Boreal Peatland Ecosystems; Ecological Studies; Springer: Berlin/Heideberg, Germany, 2006; Volume 188, pp. 1–448. [Google Scholar]
- Aerts, R.; Ludwig, F. Water-table changes and nutritional status affect trace gas emissions from laboratory columns of peatland soils. Soil Biol. Biochem. 1997, 29, 1691–1698. [Google Scholar] [CrossRef]
- Gustavsen, H.G.; Päivänen, J. Luonnontilaisten soiden puustot kasvullisella metsämaalla 1950-Luvun alussa (Tree stands on virgin forested mires in the early 1950’s in Finland). Folia For. 1986, 673, 1–27. [Google Scholar]
- Hogg, P.; Squires, P.; Fitter, A.H. Acidification, nitrogen deposition and rapid vegetational change in a small valley mire in Yorkshire. Biol. Conserv. 1995, 71, 143–153. [Google Scholar] [CrossRef]
- Limpens, J. Prospects for Sphagnum Bogs Subject to High Nitrogen Deposition. Ph.D. Thesis, Wageningen University, Wageningen, The Netherlands, 2003. Available online: https://library.wur.nl/WebQuery/wurpubs/fulltext/121392 (accessed on 5 March 2019).
- Hayward, P.M.; Clymo, R.S. The Growth of Sphagnum: Experiments on, and Simulation of, Some Effect of Light Flux and Water-Table Depth. J. Ecol. 1983, 71, 845–863. Available online: https://www.jstor.org/stable/2259597?read-now=1&seq=2#page_scan_tab_contents (accessed on 5 March 2019). [CrossRef]
- Heijmans, M.M.P.D.; Berendse, F.; Arp, W.J.; Masselink, A.K.; Klees, H.; De Visser, W.; Van Breemen, N. Effects of Elevated Carbon Dioxide and Increased Nitrogen Deposition on Bog Vegetation in the Netherlands. J. Ecol. 2001, 89, 268–279. [Google Scholar] [CrossRef]
- Tomassen, H.B.M.; Smolders, A.J.P.; Lamers, L.P.M.; Roelofs, J.G.M. Stimulated growth of Betula pubescens and Molinia caerulea on ombrotrophic bogs: Role of high levels of atmospheric nitrogen deposition. J. Ecol. 2003, 91, 357–370. [Google Scholar] [CrossRef]
- Takagi, K.; Tsuboya, T.; Takahashi, H.; Inoue, T. Effect of the invasion of vascular plants on heat and water balance in the Sarobetsu mire, Northern Japan. Wetlands 1999, 19, 246–254. [Google Scholar] [CrossRef]
- Arnolds, E. The analysis and classification of fungal communities with special reference to macrofungi. In Fungi in Vegetation Science; Winnterhoff, W., Ed.; Kluver Academic Publishers: Dordrecht, The Netherlands, 1992; pp. 7–47. [Google Scholar]
- Thormann, M.N. Diversity and function of fungi in peatlands: A carbon cycling perspective. Can. J. Soil Sci. 2006, 86, 281–293. [Google Scholar] [CrossRef]
- Thormann, M.N. The role of fungi in decomposition dynamics in peatlands. In Boreal Peatland Ecosystems; Wieder, R.K., Vitt, D.H., Eds.; Ecological Studies; Springer: Berlin, Heideberg, Germany, 2006; Volume 188, pp. 101–123. Available online: https://link.springer.com/content/pdf/10.1007%2F978-3-540-31913-9_6.pdf (accessed on 20 February 2019).
- Bujakiewicz, A. Share of macromycetes in plant communities found on the peat substrate in the Słowiński National Park. Poznań. Bad. Fizjogr. Pol. Zach. B 1986, 37, 101–129. (In Polish) [Google Scholar]
- Friedrich, S. Macromycetes in the plant associations of the Goleniowska Woods. Acta Mycol. 1986, 21, 43–76. (In Polish) [Google Scholar] [CrossRef]
- Tedersoo, L.; Bahram, M.; Toots, M.; Diedhiou, A.G.; Henkel, T.W.; Kjoller, R.; Morris, M.H.; Nara, K.; Nouhra, E.; Peay, K.G.; et al. Towards global patterns in the diversity and community structure of ectomycorrhizal fungi. Mol. Ecol. 2012, 21, 4160–4170. [Google Scholar] [CrossRef] [PubMed]
- Watling, R. Assessment of fungal diversity: Macromycetes, the problems. Can. J. Bot. 1995, 73 (Suppl. 1), S15–S24. [Google Scholar] [CrossRef]
- Ferris, R.; Peace, A.J.; Newton, A.C. Macrofungal communities of lowland Scots pine (Pinus sylvestris L.) and Norway spruce (Picea abies (L.) Karsten) plantations in England: Relationships with site factors and stand structure. For. Ecol. Manag. 2000, 131, 255–267. [Google Scholar] [CrossRef]
- Kranabetter, J.M.; Friesen, J.; Gamiet, S.; Kroeger, P. Epigeous Fruiting Bodies of Ectomycorrhizal Fungi as Indicators of Soil Fertility and Associated Nitrogen Status of Boreal Forests. Mycorrhiza 2009, 19, 535–548. Available online: https://link.springer.com/article/10.1007/s00572-009-0255-0 (accessed on 3 March 2019). [CrossRef] [PubMed]
- Rudawska, M. Studies on the Factors Regulating the Mycorrhizal Symbiosis of Pine Seedlings; Instytut Dendrologii PAN: Kórnik, Poland, 1998; pp. 1–55. (In Polish) [Google Scholar]
- Smith, S.E.; Read, D.J. Mycorrhizal Symbiosis, 2nd ed.; Academic Press: San Diego, CA, USA, 1997. [Google Scholar]
- Baar, J.; Van Groenendael, J.M.; Roelofs, J.G.M. Are Ectomycorrhizal Fungi Associated with Alnus of Importance for Forest Development in Wet Environments? Plant Biol. 2000, 2, 505–511. Available online: https://www.thieme-connect.com/products/ejournals/html/10.1055/s-2000-7468 (accessed on 8 March 2019). [CrossRef]
- Limpens, J.; Berendse, F.; Klees, H. How Phosphorus Availability Affects the Impact of Nitrogen Deposition on Sphagnum and Vascular Plants in Bogs. Ecosystems 2004, 7, 793–804. Available online: https://link.springer.com/article/10.1007/s10021-004-0274-9 (accessed on 8 February 2019). [CrossRef]
Figure 1.
Dendrogram of cluster analysis on the basis of soils parameters. Abbreviations: wet (R1, R2, R3, ZB3, N2, N3 and SD2), wetter (S1, S2, T1, ZB1, ZB2, N1, SD1 and Z1), other abbreviations are as in Table 2.
Figure 1.
Dendrogram of cluster analysis on the basis of soils parameters. Abbreviations: wet (R1, R2, R3, ZB3, N2, N3 and SD2), wetter (S1, S2, T1, ZB1, ZB2, N1, SD1 and Z1), other abbreviations are as in Table 2.

Figure 2.
Number of plant species in the patches.

Figure 3.
The vegetation cover in layers of the patches. Abbreviations: ‘a+b’—cover of tree and shrub layer, c—cover of herb layer, d—cover of moss layer.
Figure 3.
The vegetation cover in layers of the patches. Abbreviations: ‘a+b’—cover of tree and shrub layer, c—cover of herb layer, d—cover of moss layer.

Figure 4.
Contribution of bioecological groups of fungi in the patches. Abbreviations: M—mycorrhizal fungi, Sh—fungi on peat, Sm—fungi among mosses, Sl—litter-inhabiting fungi.
Figure 4.
Contribution of bioecological groups of fungi in the patches. Abbreviations: M—mycorrhizal fungi, Sh—fungi on peat, Sm—fungi among mosses, Sl—litter-inhabiting fungi.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Characteristics of the analyzed bogs.
| Bog Name | Geographical Coordinates | Area: Bog/Reserve (ha) | Dominant Communities | Kind of Protection |
|---|---|---|---|---|
| Roby | 54°6′33.6″ N 15°18′53.8″ E | 96.25/84.40 | Birch bog, willow thickets, community with Myrica gale and Erica tetralix, moss community, high sedge | Reserve; Natura 2000 PLH320017, PLB320010 |
| Stramniczka | 54°9′1.6″ N 15°41′25.2″ E | 94.49 | Boggy coniferous forests, birch bog, moss community | Reserve; Natura 2000 PLH320017 |
| Ziemomyśl | 53°11′33.7″ N 15°18′39.7″ E | 0.76 | Birch bog, community of Eriophorum vaginatum-Sphagnum fallax | – |
| Zielone Bagna | 53°40′23.7″ N 16°5′10.3″ E | 55.38 | Boggy coniferous forests, birch bog, willow thickets, moss community | Reserve; Natura 2000 PLH320039, PLB320019 |
| Mszar near Stara Dobrzyca * | 53°48′9.3″ N 15°31′58.0″ E | 11.17 | Boggy coniferous forests, birch bog, moss community | Reserve; Natura 2000 PLH320049 |
| Niewiadowo * | 53°39′25.20″ N 14°53′24.89″ E | 42 | Boggy coniferous forests, birch bog, moss community | Natura 2000 PLH320013 |
| Torfowisko Toporzyk | 53°42′40.2″ N 16°3′3.8″ E | 43.07 | Boggy coniferous forests, birch bog, moss community | Reserve; Natura 2000 PLB320019 |
*—mid-forest bogs; without stars—mid-field bogs.
Table 2.
Characteristics of the research plots.
| Bog Name | Starting Peat Extraction | Plot | Localization | Comments |
|---|---|---|---|---|
| Niewiadowo | before 1929 1 | N1 | middle part of the bog | secondary habitat—former peat extraction pit |
| N2 | middle part of the bog | habitat drying * | ||
| N3 | middle part of the bog | habitat drying *, secondary—former peat extraction pit | ||
| Roby | before 1929 2 | R1 | middle part of the bog | habitat drying *, about 30 m from the former peat extraction pit |
| R2 | middle part of the bog | habitat drying *, about 20 m from the former peat extraction pit | ||
| R3 | middle part of the bog | habitat drying * | ||
| Stramniczka | before 1929 3 | S1 | middle part of the bog | plot in the immediate vicinity of overgrowing former extraction with a moss community |
| S2 | middle part of the bog | secondary habitat—former peat extraction pit | ||
| Mszar near Stara Dobrzyca | – | SD1 | bog margin | plot on the bog, not drained and not exploited, in the depression of the area, natural habitat |
| SD2 | bog margin | plot on the bog, not drained and not exploited, less hydrated, natural habitat | ||
| Torfowisko Toporzyk | before 1921 4 | T1 | bog margin | plot in the depression of the area, exposed to surface runoff from the surrounding arable fields |
| Ziemomyśl | – | Z1 | bog margin | plot on the bog, not drained and not exploited, in the depression of the area, exposed to surface runoff from the surrounding arable fields |
| Zielone Bagna | before 1921 4 | ZB1 | middle part of the bog | secondary habitat—former peat extraction pit |
| ZB2 | middle part of the bog | secondary habitat—former peat extraction pit | ||
| ZB3 | middle part of the bog | habitat drying *, secondary—former peat extraction pit |
Table 3.
Humidity and selected chemical properties of the top layer of peat soils.
| Plot | N-NH4 mg/dm3 | N-NO3 mg/dm3 | N-NO2 mg/dm3 | P-PO4 mg/dm3 | pHH2O | pHKCl | Humidity % |
|---|---|---|---|---|---|---|---|
| Peat bog—Stramniczka | |||||||
| S1 | 3.25 abc | 2.63 ab | 0.062 abc | 1.60 ab | 4.16 cd | 2.95 ab | 86 abcd |
| S2 | 4.83 bcd | 2.41 ab | 0.058 abc | 1.36 a | 3.89 abc | 2.84 ab | 88 bcde |
| Peat bog—Zielone Bagno | |||||||
| ZB1 | 2.91 ab | 2.29 ab | 0.036 ab | 1.06 a | 4.02 bc | 3.01 ab | 90 de |
| ZB2 | 5.75 de | 2.50 ab | 0.054 ab | 1.07 a | 4.03 bc | 2.97 ab | 92 e |
| ZB3 | 3.23 abc | 2.03 a | 0.075 abc | 1.22 a | 3.78 abc | 2.74 ab | 84 abcd |
| Peat bog—Roby | |||||||
| R1 | 3.60 abcd | 3.83 abc | 0.088 abc | 1.34 a | 3.74 ab | 2.82 ab | 81 a |
| R2 | 3.64 abcd | 3.78 abc | 0.085 abc | 1.33 a | 3.70 ab | 2.81 ab | 81 a |
| R3 | 3.54 abcd | 3.70 abc | 0.078 abc | 1.33 a | 3.68 ab | 2.73 ab | 81 a |
| Peat bog—Niewiadowo | |||||||
| N1 | 3.62 abcd | 4.02 bc | 0.123 c | 2.61 bc | 4.42 de | 3.03 b | 90 de |
| N2 | 1.79 a | 2.34 ab | 0.025 a | 0.80 a | 3.60 a | 2.76 ab | 80 a |
| N3 | 5.37 cd | 5.24 c | 0.057 abc | 0.65 a | 3.77 ab | 2.67 a | 83 ab |
| Peat bog—Mszar near Stara Dobrzyca | |||||||
| SD1 | 4.22 bcd | 3.80 adc | 0.036 ab | 1.19 a | 4.60 e | 3.61 c | 92 e |
| SD2 | 2.94 ab | 2.07 a | 0.079 abc | 1.329 a | 4.49 de | 3.53 c | 84 abc |
| Peat bog—Toporzyk | |||||||
| T1 | 4.22 bcd | 3.61 abc | 0.034 ab | 1.22 a | 4.57 e | 3.67 c | 88 bcde |
| Peat bog—Ziemomyśl | |||||||
| Z1 | 7.74 e | 2.98 ab | 0.095 bc | 2.80 c | 4.03 bc | 3.03 b | 89 cde |
Number of samples: N = 7; different letters (a, b, c, d, e—homogeneous groups) indicate significant difference at p < 0.05.
Table 4.
Characteristics of birch bog habitats varied in terms of humidity.
| Habitat | Humidity % | pHH2O | pHKCl | N-NO3 mg/dm3 | N-NO2 mg/dm3 | N-NH4 mg/dm3 | P-PO4 mg/dm3 |
|---|---|---|---|---|---|---|---|
| Wet | 82.08 b | 3.82 b | 2.86 b | 3.28 | 0.070 | 3.44 b | 1.14 b |
| Wetter | 89.51 a | 4.21 a | 3.14 a | 3.03 | 0.077 | 4.57 a | 1.61 a |
Number of samples: Nwet = 49 and Nwetter = 56; different letters (a, b) indicate significant difference at p < 0.05.
Table 5.
Characteristics of peat soil research areas on natural habitats (SD1-2) subjected to surface runoffs (T1 and Z1) and not subjected to surface runoff (S1-2, R1-3, ZB1-3, N1-3 and Z1).
Table 5.
Characteristics of peat soil research areas on natural habitats (SD1-2) subjected to surface runoffs (T1 and Z1) and not subjected to surface runoff (S1-2, R1-3, ZB1-3, N1-3 and Z1).
| Parameter | Humidity % | pHH2O | pHKCl | N-NO3 mg/dm3 | N-NO2 mg/dm3 | N-NH4 mg/dm3 | P-PO4 mg/dm3 |
|---|---|---|---|---|---|---|---|
| Surface runoff | 88.87 a | 4.30 a | 3.35 b | 3.30 | 0.064 | 5.60 a | 2.01 a |
| Natural | 88.12 a | 4.54 a | 3.57 a | 2.93 | 0.058 | 3.58 b | 1.26 b |
| Without surface runoff | 85.15 b | 3.88 b | 2.84 c | 3.15 | 0.080 | 3.77 b | 1.31 b |
Number of samples: N surface runoff = 14, N natural = 14 and N without surface runoff = 77; different letters (a, b, c) indicate significant difference at p < 0.05.
Table 6.
Characteristics of peat soil habitats depending on their location on the bog. Former peat extraction pits (S1-2, ZB1-3, N1 and N3), bog margin (T1, SD1-2 and Z1), middle part of the bog (R1-3 and N2).
Table 6.
Characteristics of peat soil habitats depending on their location on the bog. Former peat extraction pits (S1-2, ZB1-3, N1 and N3), bog margin (T1, SD1-2 and Z1), middle part of the bog (R1-3 and N2).
| Plot Location | Humidity % | pHH2O | pHKCl | N-NO3 mg/dm3 | N-NO2 mg/dm3 | N-NH4 mg/dm3 | P-PO4 mg/dm3 |
|---|---|---|---|---|---|---|---|
| Peat extraction pits | 87.59 a | 4.01 b | 2.89 b | 3.01 | 0.083 | 4.14 ab | 1.37 |
| Bog margin | 88.50 a | 4.22 a | 3.46 a | 3.11 | 0.061 | 4.78 a | 1.63 |
| Middle part of the bog | 80.89 b | 3.68 c | 2.77 b | 3.41 | 0.069 | 3.14 b | 1.20 |
Number of samples: N peat extraction pits = 49, N bog margin = 28 and N middle part of the bog = 28; different letters (a, b, c) indicate significant difference at p < 0.05.
Table 7.
Characteristics of peat soils of mid-field and mid-forest bogs. Mid-forest bogs (N1-3 and SD1-2), mid-field bogs (S1-2, T1, R1-3, ZB1-3 and Z1).
Table 7.
Characteristics of peat soils of mid-field and mid-forest bogs. Mid-forest bogs (N1-3 and SD1-2), mid-field bogs (S1-2, T1, R1-3, ZB1-3 and Z1).
| Parameter | Humidity % | pHH2O | pHKCl | N-NO3 mg/dm3 | N-NO2 mg/dm3 | N-NH4 mg/dm3 | P-PO4 mg/dm3 |
|---|---|---|---|---|---|---|---|
| Mid-field bogs | 85.86 | 3.96 b | 2.96 | 2.97 | 0.066 | 4.27 | 1.43 |
| Mid-forest bogs | 86.14 | 4.18 a | 3.12 | 3.49 | 0.087 | 3.59 | 1.31 |
Number of samples: N mid-forest bogs = 35 and N mid-field bogs = 70; different letters (a, b) indicate significant difference at p < 0.05.
Table 8.
Pearson correlation (R - coefficient) between plants, macromycetes and habitat parameters. Abbreviations: NP—total number of plant specis in relevé, Vac-Pic—species of Vaccinio-Picetea, Ox-Sph— species of Oxycocco-Sphagnetea, a + b—cover of tree and shrub layer (%), c—cover of herb layer (%), d—cover of moss layer (%), NF—total of fungi species, M—mycorrhizal fungi, Sh—fungi on peat, Sm—fungi among mosses, Sl—fungi on litter.
Table 8.
Pearson correlation (R - coefficient) between plants, macromycetes and habitat parameters. Abbreviations: NP—total number of plant specis in relevé, Vac-Pic—species of Vaccinio-Picetea, Ox-Sph— species of Oxycocco-Sphagnetea, a + b—cover of tree and shrub layer (%), c—cover of herb layer (%), d—cover of moss layer (%), NF—total of fungi species, M—mycorrhizal fungi, Sh—fungi on peat, Sm—fungi among mosses, Sl—fungi on litter.
| Parameter | Humidity | pHH2O | pHKCl | P-PO4 | N-NH4 | N-NO3 | N-NO2 |
|---|---|---|---|---|---|---|---|
| NP | 0.60 * | 0.31 | 0.29 | 0.33 | 0.35 | 0.35 | 0.35 |
| Vac-Pic | 0.52 * | 0.49 | 0.82 * | 0.73 * | 0.72 * | 0.68 * | 0.72 * |
| Ox-Sph | −0.42 | −0.30 | −0.53 * | −0.44 | −0.48 | −0.43 | −0.48 |
| a + b | 0.56 * | 0.51 * | 0.42 | 0.45 | 0.47 | 0.47 | 0.47 |
| c | 0.54 * | −0.12 | 0.24 | 0.21 | 0.25 | 0.20 | 0.25 |
| d | 0.19 | 0.56 * | −0.17 | 0.02 | 0.09 | 0.18 | 0.09 |
| NF | 0.57 * | 0.48 | 0.94 * | 0.83 * | 0.80 * | 0.73 * | 0.80 * |
| M | 0.64 * | 0.67 * | 0.92 * | 0.92 * | 0.84 * | 0.83 * | 0.84 * |
| Sh | 0.44 | 0.19 | 0.73 * | 0.58 * | 0.58 * | 0.48 | 0.58 * |
| Sm | −0.24 | 0.01 | −0.41 | −0.26 | −0.23 | −0.14 | −0.23 |
| Sl | 0.50 | 0.39 | 0.82 * | 0.73 * | 0.73 * | 0.67 * | 0.73 * |
0.60 *—significant by p < 0.05, N = 15.
Table 9.
Pearson correlation (R-coefficient) between plant predictors and macromycetes.
| Parameter | NP | Vac-Pic | Ox-Sph | a + b | c | d |
|---|---|---|---|---|---|---|
| NF | 0.21 | 0.87 * | −0.60 * | 0.34 | 0.25 | −0.43 |
| M | 0.24 | 0.67 * | −0.30 | 0.37 | 0.11 | −0.14 |
| Sh | 0.19 | 0.72 * | −0.61 * | 0.22 | 0.43 | −0.59 * |
| Sm | −0.31 | −0.49 | 0.44 | −0.40 | −0.39 | 0.50 |
| Sl | 0.05 | 0.78 * | −0.46 | 0.26 | 0.27 | −0.42 |
0.87 *—significant by p < 0.05, N = 15.
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Sotek, Z.; Stasińska, M.; Malinowski, R.; Gamrat, R.; Gałczyńska, M. Birch Bog on Anthropogenically Transformed Raised Bogs. A Case Study from Pomerania (Poland). Water 2019, 11, 1224. https://doi.org/10.3390/w11061224
AMA Style
Sotek Z, Stasińska M, Malinowski R, Gamrat R, Gałczyńska M. Birch Bog on Anthropogenically Transformed Raised Bogs. A Case Study from Pomerania (Poland). Water. 2019; 11(6):1224. https://doi.org/10.3390/w11061224
Chicago/Turabian StyleSotek, Zofia, Małgorzata Stasińska, Ryszard Malinowski, Renata Gamrat, and Małgorzata Gałczyńska. 2019. "Birch Bog on Anthropogenically Transformed Raised Bogs. A Case Study from Pomerania (Poland)" Water 11, no. 6: 1224. https://doi.org/10.3390/w11061224
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.